Основной продукт азотного обмена мочевина у тритона. Азотистый обмен
Азотистый обмен - совокупность химических превращений азотсодержащих веществ в организме.
Азотистый обмен включает обмен простых и сложных белков, нуклеиновых кислот, продуктов их распада (пептидов, аминокислот и нуклеотидов), содержащих азот жироподобных веществ (липидов), аминосахаров, гормонов, витаминов и другое.
Для нормального течения процессов жизнедеятельности организм должен быть обеспечен необходимым количеством усвояемого азота. Главнейшей составной частью и основным источником азота пищи человека являются белковые вещества (см. Белки).
Суточная норма белка в питании взрослого человека, принятая в СССР, составляет 100 г белка или 16 г белкового азота при трате энергии в 2500 ккал. Однако и значительно меньшие количества белка могут обеспечить азотистое равновесие, то есть состояние, при котором количества вводимого и выводимого азота одинаковы. После приема белковой пищи основной обмен (см. Обмен веществ и энергии) повышается больше, чем это обусловлено калорийной ценностью белка. Это явление получило наименование «специфически динамическое действие» белковой пищи. Механизм этого явления не вполне ясен. По-видимому, некоторые аминокислоты - продукты расщепления белка - участвуют в реакциях, связанных с гидролизом АТФ и образованием АДФ, обусловливая повышенное потребление кислорода.
При общем голодании или при недостаточном азотистом питании количество выводимого с мочой и калом азота превышает количество вводимого с пищей - состояние отрицательного азотистого баланса. В том случае, когда количество вводимого азота превышает количество выводимого, наступает состояние положительного азотистого баланса, что характерно для растущего организма, при процессах регенерации и так далее.
При составлении или оценке рациона питания необходимо учитывать полноценность белков, характеризующуюся содержанием в них незаменимых аминокислот, то есть таких, которые не могут образоваться из других соединений в организме (см. Аминокислоты).
Суточная потребность организма человека в различных незаменимых аминокислотах неодинакова (табл. 1).
Таблица 1. Потребность взрослого человека в незаменимых аминокислотах (в г за сутки)
Переваривание белков и других азотсодержащих веществ
Для высокоорганизованных позвоночных животных, в том числе и для человека, началом процессов азотистого обмена следует считать переваривание в желудочно-кишечном тракте простых и сложных белков, а также других сложных азотистых соединений с последующим всасыванием продуктов их расщепления.
Переваривание белков начинается в желудке под влиянием ферментов пепсина (см.) и гастриксина, вырабатываемых в слизистой оболочке желудка в неактивной форме - в виде зимогенов (проферментов).
Кислая среда, необходимая для активации зимогенов, обеспечивается соляной кислотой, секретируемой железами слизистой оболочки дна желудка (обкладочные клетки). Пепсин (оптимум рН около 2) и гастриксин (оптимум рН 3-4) представляют собой протеазы - эндопептидазы, которые разрывают пептидные связи между аминокислотами, расположенными внутри пептидных цепей молекулы белка (см. Пептидгидролазы).
При накоплении в желудке пищевой массы, обладающей достаточно кислой реакцией, рефлекторно раскрывается пилорический жом, и пищевая масса порциями поступает в двенадцатиперстную кишку, а затем в нижележащие отделы тонкой кишки, где в дальнейшем расщеплении пептидных связей участвуют ферменты сока поджелудочной железы - трипсин (см.), химотрипсин (см.) и карбоксипептидаза (см.) и ферменты кишечника - амино- и дипептидазы.
Трипсин и химотрипсин относятся к эндопептидазам (оптимум рН около 8,0), карбокси- и аминопептидазы - к экзопептидазам; они расщепляют крайнюю пептидную связь соответственно со стороны свободной карбоксильной и аминогруппы. При образовании дипептидов они расщепляются дипептидазами. Параллельно перевариванию простых белков в тонкой кишке происходит расщепление нуклеопротеидов, а также дезоксирибонуклеиновых (ДНК) и рибонуклеиновых (РНК) кислот.
В результате последовательного проявления гидролитической активности ферментов пищеварительных желез, а также микроорганизмов кишечника простые и сложные белки, а также другие биополимеры распадаются, и продукты распада (низкомолекулярные пептиды, аминокислоты, нуклеотиды, нуклеозиды) всасываются в тонком кишечнике и поступают в кровь.
Параллельно происходит ферментативный распад тканевых белков под влиянием тканевых протеаз - катепсинов (см.) и пептидаз (см. Пептидгидролазы). Образующиеся ори этом продукты распада также попадают в кровь и разносятся во все органы и ткани.
Тканевой обмен аминокислот
Фонд аминокислот, образовавшийся в результате ферментативного расщепления пищевых продуктов или продуктов распада тканей, расходуется на биосинтез белков и многих других соединений, свойственных только данному организму, на энергетические затраты, а также на образование конечных продуктов азотистого обмена, подлежащих выведению (рис. 1).
Участие аминокислот в процессах биосинтеза
Синтез специфических для данного организма белков находится под контролем молекул ДНК, входящих в состав хроматина клеточных ядер.
На одном из тяжей ДНК (в месте ее раскручивания) по закону комплементарности (см. Генетический код) происходит сборка (синтез) информационных, или матричных, РНК (мРНК).
К фиксированным на рибосомах мРНК подходят транспортные рибонуклеиновые кислоты (тРНК), несущие на себе предварительно активированные аминокислоты, которые фиксируются на мРНК. Рядом располагаются такие аминокислоты, которые в синтезируемом белке должны быть соединены пептидной связью, чем обеспечивается специфическая первичная структура белков со строго определенным порядком следующих друг за другом аминокислот.
В свою очередь первичная структура предопределяет, если не полностью, то в значительной мере, пространственную конфигурацию, или третичную структуру, белков, в том числе и белков-ферментов.
Выпадение или нарушение какого-либо звена в сложном процессе биосинтеза фермента, осуществляющего определенную реакцию в обмене веществ, может привести к тяжелым патологическим нарушениям. Так, причиной многих наследственных болезней (см.) является выпадение синтеза всего лишь одного белка-фермента (например, гидроксилазы при фенилпировиноградной олигофрении); «ошибка» в первичной структуре У α- или β-цепей гемоглобина, заключающаяся в замене всего лишь одной из 287 аминокислот, приводит к образованию патологических форм гемоглобина с нарушенной функцией присоединения и отдачи кислорода.
Фонд аминокислот используется также при синтезе других соединений.
Например, биосинтез пуриновых нуклеотидов (см. Пуриновые основания), начинающийся с рибозил-5-фосфата, проходит через многочисленные стадии и завершается образованием инозиновой кислоты (инозиновая кислота затем может подвергаться превращениям в адениловую и гуаниловую кислоты). При этом требуется участие глутамина (амида глутаминовой кислоты) в качестве источника азота в 3-м и 9-м положениях, глицина - в 7-м положении и углерода - в 4-м и 5-м положениях. Аспарагиновая к-та - источник азота в 1-м положении:


При последующем образовании адениловой кислоты (см. Аденозинфосфорные кислоты) вновь вовлекается аспарагиновая кислота, азот которой обеспечивает аминогруппу, стоящую при 6-м углеродном атоме пуринового кольца. При синтезе гуаниловой кислоты (см.) аминогруппа при 2-м углеродном атоме берется из глутамина.

из аммиака (NH3), бикарбоната (НСО3-), аденозинтрифосфата (АТФ) как источника энергии и, наконец, N-ацетилглутаминовой кислоты в качестве активатора:
Карбамильная группа карбамилфосфата ферментативно переносится на аспарагиновую кислоту. Через образовавшуюся карбамиласпарагиновую к-ту, дигидрооротовую и оротовую кислоты (рис. 2) образуется оротидиловая кислота, переходящая в уридиловую кислоту и уридинтрифосфат (УТФ). Путем аминирования УТФ образуется цитидинтрифосфат (ЦТФ), причем эта последняя реакция представляет собой регулируемый процесс по закону обратной связи: ЦТФ тормозит образование карбамиласпарагиновой кислоты, а АТФ снимает это торможение. Таким образом, образование пиримидиновых нуклеотидов, входящих в состав нуклеиновых кислот, регулируется соотношением содержания ЦТФ и АТФ.

Помимо образования пуриновых и пиримидиновых нуклеотидов, аминокислоты участвуют в образовании многих других физиологически важных соединений.
 в результате ряда последовательных ферментативных превращений образуется никотиновая кислота, выполняющая функцию антипеллагрического витамина и участвующая в виде никотинамида
в результате ряда последовательных ферментативных превращений образуется никотиновая кислота, выполняющая функцию антипеллагрического витамина и участвующая в виде никотинамида
 в биосинтезе никотпнамидных коферментов НАД и НАДФ.
в биосинтезе никотпнамидных коферментов НАД и НАДФ.
2. Простейшая аминокислота глицин (CH 2 NH 2 COOH), помимо участия в образовании пуринов, обеспечивает весь азот и ряд атомов углерода при биосинтезе порфиринов, составляющих структурную основу желчных пигментов и небелковой части (простетической группы) железосодержащих хромопротеидов (см.).
Глицин выполняет также роль акцептора амндиновой группы аргинина при синтезе гуанидинуксусной кислоты, N-метилпронзводное которой - креатин (см.)
 является важной составной -частью скелетной мускулатуры, сердца и мозга, а в виде фосфорилированного продукта (фосфокреатина) обеспечивает резерв богатых энергией фосфорных соединений, необходимых для функциональной активности ткани.
является важной составной -частью скелетной мускулатуры, сердца и мозга, а в виде фосфорилированного продукта (фосфокреатина) обеспечивает резерв богатых энергией фосфорных соединений, необходимых для функциональной активности ткани.
3. Серин участвует в образовании сложного аминоспирта - сфингозина (см. Сфингозины), входящего в состав сфингомиелина (см. Сфинголипиды) - липида, особенно богато представленного в составе мозга и нервной ткани.
 Серин участвует также в синтезе кофермента (см.) ацетилирования (КоА), ацилпроизводные которого представляют активную форму жирных кислот (см. Жировой обмен), участвующих в различных процессах биосинтеза и окислительного распада.
Серин участвует также в синтезе кофермента (см.) ацетилирования (КоА), ацилпроизводные которого представляют активную форму жирных кислот (см. Жировой обмен), участвующих в различных процессах биосинтеза и окислительного распада.
Таблица 2. Некоторые биологически важные азотистые вещества, образующиеся из аминокислот
В табл. 2 представлены дополнительные сведения об отдельных аминокислотах, являющихся предшественниками некоторых других биологически важных азотистых соединений.
Функциональные группы аминокислот широко вовлекаются в различные реакции обмена веществ.
Прежде всего это относится к аминогруппам, участвующим в реакции переаминирования (см.). Эта реакция, представляющая важнейший путь ферментативного превращения аминокислот, была открыта советскими биохимиками А. Е. Браунштейном и М. Г. Крицман в 1937 году. Она заключается в обратимом ферментативном переносе α-аминогруппы α-аминокислоты на α-углеродный атом α-кетокислоты без промежуточного освобождения аммиака.
В реакциях переаминирования, катализируемых различными трансаминазами, могут участвовать не только аминогруппы α-аминокислот, но и аминогруппы аминов и ω-аминокислот (например, β-аланина, γ-аминомасляной кислоты); акцептировать аминогруппы могут не только α-кетокислоты, но и альдегиды (например, малоновый или янтарный полуальдегиды).
Общая схема реакции переаминирования обычно изображается в следующем виде:
Непременным участником обратимой реакции ферментативного переаминирования, выполняющим коферментную функцию, является пиридоксальфосфат (I), а также пиридоксаминфосфат (II), оба - производные витамина В6 (пиридоксина).
 Пиридоксальфосфат принимает на себя аминогруппу аминокислоты и через стадии образования шиффовых оснований превращается в пиридоксаминфосфат (II), который передает аминогруппу также через промежуточные стадии на кетокислоту, возвращаясь в первоначальное состояние (I).
Пиридоксальфосфат принимает на себя аминогруппу аминокислоты и через стадии образования шиффовых оснований превращается в пиридоксаминфосфат (II), который передает аминогруппу также через промежуточные стадии на кетокислоту, возвращаясь в первоначальное состояние (I).
Дикарбоновые аминокислоты - глутаминовая и аспарагиновая - наиболее активные участники процесса переаминирования. Под влиянием фермента глутаматдегидрогеназы осуществляется образование глутаминовой кислоты из аммиака и кетоглутаровой кислоты. Аминогруппа глутаминовой кислоты широко транспортируется при участии аминофераз на различные α-кетокислоты и альдегиды, образуя новые аминокислоты и амины. Этим косвенным путем азот аммиака вовлекается в состав многочисленных азотистых органических веществ.
В биосинтезе ряда биологически активных соединений значительная роль принадлежит процессу метилирования. Перенос метильной группы, как правило, осуществляется аминокислотой - метионином в виде аденозилметионина, превращающегося после отдачи метильной группы в S-аденозилгомоцистеин (рис. 3).

Акцепторы метильной группы разнообразны; к ним относятся: липиды, транспортные нуклеиновые кислоты, содержащие минорные (редкие) компоненты - метилированные нуклеотиды, гуанидинуксусную кислоту, никотинамид и другое. Донорами метильных групп могут быть, помимо аденозилметионина, еще и холин, бетаины, N5-метилтетрагидрофолиевая кислота и другое. (см. Метилирование).
Участие аминокислот в процессах катаболизма
Одним из путей катаболизма (деградации) аминокислот является их ферментативное декарбоксилирование (см.), приводящее к освобождению углекислого газа и образованию биогенных аминов, обладающих высокой биологической активностью, например гистамина
 из гистидина; серотонина
из гистидина; серотонина
 из окситриптофана; γ-аминомасляной к-ты из глутаминовой к-ты:
из окситриптофана; γ-аминомасляной к-ты из глутаминовой к-ты:
H 2 N-CH 2 -CH 2 -CH 2 -COOH
тирамина из тирозина

Декарбоксилирование аминокислот катализируется декарбоксилазами (см.), коферментом которых обычно является пиридоксальфосфат, однако механизм декарбоксилирования аминокислот остается недостаточно выясненным. В декарбоксилазе гистидина коферментная функция принадлежит остатку пировиноградной кислоты, карбоксильная группа которой соединена с пептидной цепью белка-фермента кислотноамидной связью:

Образовавшиеся при декарбоксилировании аминокислот амины служат субстратами окисления для моноамино- и диаминооксидаз - ферментов, отличающихся друг от друга не только белковой частью, но и коферментами: митохондриалыше моноаминооксидазы (см.) принадлежат к флавопротеидам, их кофсрмент - флавинадениндинуклеотид. У диаминооксидаз коферментом служит пиридоксальфосфат. Образующиеся при дезаминированип монаминов аммиак и альдегиды претерпевают дальнейшие превращения: обезвреживание аммиака происходит преимущественно путем образования мочевины (см.), углеродный скелет аминов (в виде альдегидов) подвергается дальнейшему окислению.
Другим процессом деградации L-аминокислот является их окислительное дезаминирование (см.), идущее с образованием аммиака и кетокислот. Эта реакция протекает в организме высших животных и человека очень медленно (в противоположность окислительному образованию аммиака из D-аминокислот), однако может осуществляться быстрее косвенным путем: сначала при персаминировании образуется α-глутаминовая кислота, которая затем при дезаминировании является источником кетоглутаровой кислоты и аммиака. Следует, однако, учесть, что в реакции дезаминирования равновесие смещено в сторону восстановительного образования глутаминовой кислоты, то есть слева направо:
Пути образования аммиака из аминокислот остаются недостаточно ясными.
В последнее время Г. X. Бунатяном и его сотрудниками большое значение в процессе образования аммиака (в частности, в центральной нервной системе и в печени) приписывается отщеплению NH2-группы аденина, находящегося в составе никотинамидадениндпнуклеотида (НАД). Продуктом этой реакции является дезамгшоникотинамид-адениндинуклеотид (деНАД):

Последующее аминирование деНАД осуществляется при участии аспарагиновой кислоты, протекает с образованием промежуточного продукта (НАД-янтарной кислоты) и после отщепления фумаровой кислоты приводит к восстановлению первоначальной структуры НАД (рис. 4):

Следовательно, по Г. X. Бунатяну, дезаминирование α-аминокислот с образованием аммиака протекает через образование аспарагиновой кислоты путем переаминирования, передачи аминогруппы на деНАД, образования НАД и отщепления аммиака от НАД.
В настоящее время еще невозможно оценить, насколько широко этот процесс представлен в организме и каково его биологическое значение. ДеНАД приписывают высокую биологическую активность в качестве фактора, легко проникающего как в окисленной, так и в восстановленной форме через мембрану митохондрий и значительно повышающего энергетическую эффективность окислительного фосфорилирования.
Образование конечных продуктов обмена простых белков
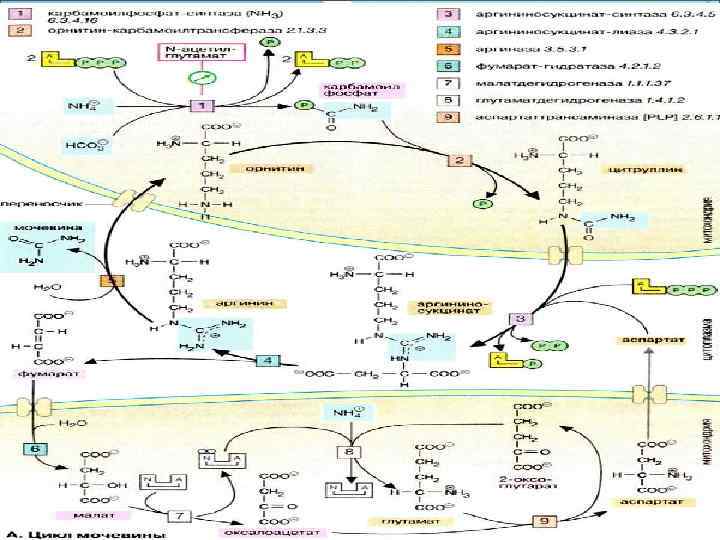
Возникший в процессах обмена веществ аммиак и безазотистый остаток аминокислот претерпевают своеобразные превращения. Основной путь нейтрализации и связывания аммиака у уреотелических животных заключается в синтезе мочевины, протекающем в печени и состоящем из серии последовательных ферментативных реакций. Первый этап этого процесса заключается в образовании карбамилфосфата (так же, как при синтезе пиримидиновых оснований), затем карбамильная группа акцептируется орнитином.
Образовавшийся при этом цитруллин эндергонически реагирует с аспарагиновой кислотой. Образовавшаяся аргининянтарная кислота подвергается расщеплению: один из продуктов реакции - фумаровая кислота - включается в цикл трикарбоновых кислот, другой - аргинин - гидролитически расщепляется аргиназой на мочевину и орнитин (рис. 5). Последний вновь включается в цепь превращений, приводящих к образованию мочевины. Процесс этот, получивший наименование орнитинового цикла, протекает в печени, хотя отдельные его реакции представлены также в сердце, ткани мозга и другое.

Таким образом, азот, выводимый из организма в виде мочевины, наполовину берется из аммиака и наполовину из аспарагиновой кислоты.
Нейтрализация образующегося в организме аммиака происходит еще путем синтеза амидов - аспарагина и глутамина. Амидная группа последнего участвует в синтезе пуринов, нуклеиновых кислот и так далее.
Нейтрализация аммиака у урикотелических животных (рептилии, птицы) связана с образованием мочевой кислоты (см.).
Безазотистая часть аминокислот, как правило, включается через многочисленные промежуточные этапы в разные стадии окислительных превращений по циклу трикарбоновых кислот (см. Трикарбоновых кислот цикл).
Согласно схеме, приведенной на рис. 6, отчетливо выявляется роль аминокислот в обеспечении энергетических запросов организма. Нарушения в превращениях тех или иных аминокислот часто генетически обусловлены и являются причиной различных болезней.
Причиной нарушений, как правило, является дефект в одном специфически действующем ферменте или в серии ферментативных реакций. Эти нарушения могут возникнуть, например, в силу недостаточного образования или слишком быстрого расщепления кофермента, участвующего во многих ферментативных процессах.
Тканевой обмен нуклеотидов
Продукты распада нуклеопротеидов и нуклеиновых кислот - нуклеотиды и нуклеозиды - претерпевают в органах и тканях различные превращения.
Синтез ДНК и РНК
Нуклеотиды - как пуриновые, так и пиримидиновые - участвуют в синтезе нуклеиновых кислот в клеточных ядрах. Синтез ДНК осуществляется ферментами - ДНК-полимеразами, для которых субстратами служат дезоксирибонуклеозидтрифосфаты.
Синтез ДНК сопровождается освобождением молекул пирофосфата в количестве, соответствующем числу молекул нуклеозидтрифосфатов, вступивших в реакцию. ДНК (образец) и вновь синтезированный полинуклеотид образуют вместе двутяжную ДНК. Схема этого процесса может быть представлена в следующем виде:

Буква «d» перед символом нуклео-зидтрифосфата или мононуклеотидов в синтезированной молекуле ДНК обозначает, что в биосинтезе участвуют нуклеотиды, в которых пентоза представлена дезоксирибозой, то есть дезоксирибонуклеотиды. Образование дезоксирибонуклеотидов происходит в результате сложного процесса восстановления рибонуклеотидов при действии нечувствительного к нагреванию белка - тиоредоксина.
Восстановленная форма тиоредоксина образуется под действием редуктазы (фермента флавопротешговой природы), коферментом которого служит восстановленный никотинамидадениндпнуклеотидфосфат (НАДФ) по схеме:
Вновь образованная ДНК и служившая шаблоном ДНК могут на своих концах соединиться под влиянием фермента ДНК-лигазы и образовать циклическую структуру ДНК.

Синтез РНК осуществляется при участии полинуклеотидфосфорилазы - фермента, обусловливающего обратимую реакцию соединения нуклеозидднфосфатов в присутствии ионов магния и первоначальной РНК:

Образованный полимер содержит 3′-5′-фосфодиэфирные связи, которые расщепляются рибонуклеазой. Реакция обратима и может быть направлена справа налево (в сторону распада полимера) при увеличении концентрации неорганического фосфата. Первоначальная РНК в данном случае не играет роли шаблона, по которому синтезируется полинуклеотид. Скорее всего свободная ОН-группа, находящаяся в концевом нуклеотиде РНК, необходима для присоединения к ней последующих нуклеотидов независимо от входящих в их состав оснований.
По-видимому, в интактной клетке полинуклеотидфосфорилазе принадлежит функция не образования полимера, а расщепления РНК. Что касается высокополимерной РНК с определенной последовательностью нуклеотидов, то образование ее осуществляется РНК-полимеразой, действие которой аналогично ферменту, синтезирующему ДНК. РНК-полимераза активна в присутствии ДНК-шаблона, осуществляет синтез РНК из нуклеозидтрифосфатов и собирает их в последовательности, предопределенной структурой ДНК:

Катаболизм ДНК и РНК
Деградация ДНК и РНК происходит по этапам. Ферменты, расщепляющие РНК, - рибонуклеазы - широко представлены в различных животных тканях. Под влиянием рибонуклеаз двоякого типа - трансфераз и истинных гпдролаз - из РНК образуются олиго- и мононуклеотиды.
Ферменты, расщепляющие ДНК, принадлежат к нуклеазам - гидролазам; результатом их действия является образование олигонуклеотидов как с концевым 5′-фосфатом, так и 5′-фосфатом. Под влиянием диэстераз, специфических нуклеотидаз, фосфорилаз, фосфатаз и нуклеозидаз происходит деградация нуклеотидов с образованием свободных пуриновых и пиримндиновых оснований, дальнейшее превращение которых идет по разным путям.
Пуриновые основания - аденин и гуанин - подвергаются гидролитическому дезаминированию под влиянием ферментов аденазы и гуаназы. Из аденина образуется 6-оксипурин (гипоксантин), из гуанина - 2,6-диоксипурин (ксантин). Эти превращения ампнопуринов могут происходить и без предварительного распада соответствующих нуклеотидов и нуклеозидов. Гипоксантин и ксантин подвергаются дальнейшему окислению под влиянием фермента ксантиноксидазы. Конечным продуктом этого окисления является 2, 6, 8-триоксипурин, или мочевая кислота (см.). У человека мочевая кислота дальнейшим превращениям не подвергается и является постоянной составной частью мочи и конечным продуктом обмена пуриновых нуклеотидов и пуриновых оснований. У большинства млекопитающих мочевая кислота не представляет собой конечного продукта обмена веществ и переходит в аллантоин под действием фермента уриказы.

Стадии дезаминирования и окисления пуриновых оснований представлены на рис. 7.
По иному пути идет деградация пиримидиновых оснований (см.). Первый шаг заключается в восстановлении урацила в дигидроурацил с последующим гидролизом, приводящим к образованию сначала β-уреидопропионовой кислоты, а затем β-аланина, NH3 и СО2 (рис.8):

Аналогичные превращения тимина приводят к образованию β-аминоизо-масляной кислоты.
Таким обраом, конечным продуктом обмена пуринов у человека является мочевая кислота, а пиримидинов - углекислый газ и аммиак, которые могут быть источниками образования мочевины. Что касается β-аланина, то эта аминокислота участвует в биосинтезе дипептидов карнозина (см.) и ансерина (см.), в большом количестве содержащихся в скелетной мускулатуре позвоночных животных.

В мышечной ткани человека содержится лишь карнозин.
β-аланин является также составной частью пантотеновой кислоты, а следовательно, и коэнзима А, играющего очень важную роль в обмене жирных кислот, стеринов, а также в цикле трикарбоновых кислот.
Регуляция процессов азотистого обмена
Азотисный обмен так же, как и все виды обмена веществ, регулируется нервной системой как непосредственно, так и через воздействие на железы внутренней секреции. Основное значение нервной регуляции Азотистый обмен заключается в приспособлении его к изменяющимся условиям внешней и внутренней среды. В силу этого выпадение нервных воздействий на органы и ткани приводит к тяжелым нарушениям их структуры и функции.
Благодаря весьма сложным и тонким механизмам регуляции азотистого обмена у взрослого здорового человека обеспечивается относительное постоянство состава азотистых компонентов органов, тканей и внутренней среды организма. Избыток вводимых с пищей азотистых соединений выводится с мочой и калом, недостаток - пополняется из состава тканей тела (это не относится к числу незаменимых соединений).
Насколько состав крови и тканей обладает относительным динамическим постоянством, настолько изменчив состав мочи, отражающий особенности обмена веществ в значительно большей степени, чем состав плазмы крови или цельной крови. В силу этого для выводов об особенностях азотистого обмена необходимо прежде всего знать качественный и количественный состав принимаемой пищи, исследовать качественный и количественный состав азотистых соединений, выделяемых с мочой и калом, и сопоставить полученные данные. Определение особенностей состава крови может дать представление о качественном своеобразии некоторых сторон азотистого обмена, но не позволяет дать заключение о его состоянии в целом. Например, увеличение содержания белков в пищевом рационе приведет к незначительному повышению содержания небелковых азотистых составных частей (остаточного азота) крови, но выделение азотистых соединений и в первую очередь мочевины с мочой будет значительно повышено. Нарушение окислительного фосфорилирования несколько изменит соотношение содержания креатина и креатинина Р сыворотке крови, однако значительно резче изменится содержание этих соединений и их соотношение в моче (см. Креатин, Креатинурия).
Несмотря на всю сложность и разнообразие реакций, протекающих в организме, выделяемые конечные продукты обмена веществ остаются для данного вида при данном режиме питания качественно более или менее постоянными. Они претерпевают значительные отклонения от нормы при различных патологических состояниях организма.
Методические приемы изучения отдельных стадий превращения азотистых соединений значительно расширились и обогатились. Большую роль сыграло внедрение в практику исследовании азотистого обмена органических веществ, содержащих радиоактивные или тяжелые изотопы различных элементов и в первую очередь фосфора, углерода, серы, азота, кислорода, водорода. Использование этих соединений позволило довольно детально следить за их превращениями, за постепенным переходом метки из одного вещества к другому, завершающимся выделением изотопа в составе конечных продуктов обмена веществ. В настоящее время можно получить практически любую меченую аминокислоту, принимающую участие в процессах биосинтеза белков, свойственных данному организму. Эти эксперименты позволяют выяснить, где и когда аминокислоты включаются в состав белков, с какими веществами или структурами связываются аминокислоты до того, как войдут в состав пептидной цепи. Если аминокислоту глицин, меченную изотопом N 15 , ввести с пищей в организм животного, то значительная часть изотопа будет быстро выведена из организма в составе мочевины, другая часть остается в тканях и выводится очень медленно. Большая часть введенного препарата с изотопом N 15 обнаруживается в белках, причем треть меченого азота включается в белок в виде остатков глицина, а остальные две трети - в составе других аминокислотных остатков. С помощью меченых соединений были открыты или уточнены многие стадии метаболических процессов, идущих в клетках. Так, например, было подтверждено, что аминогруппы переходят от одной аминокислоты к другой (процесс переаминирования).
Патология азотистого обмена
Патология азотистого обмена проявляется в форме патологии синтеза белков и нарушений обмена различных азотсодержащих метаболитов (аминокислоты, мочевина, аммиак, креатин и креатинин, мочевая кислота и другое), циркулирующих в крови и выделяемых почками.
Основная форма патологии синтеза белков - белковая недостаточность - наступает при нарушении соотношения между процессами биосинтеза и катаболизма белковых структур, приводящем к преобладанию процессов распада над синтезом. Общая белковая недостаточность, характеризующаяся ограничением синтеза многих белков (тканевых, плазменных, ферментных), развивается при алиментарном их дефиците - при общем недоедании и при дефиците энергетических компонентов пищи - углеводов и жиров. В последнем случае белки расходуются в организме в качестве источника энергии (см. Обмен веществ и энергии). Такой же механизм развития общей белковой недостаточности имеет место при нарушении усвоения отдельных пищевых продуктов в связи с патологией пищеварительного аппарата. Ускоренная эвакуация пищи из желудка, а также гипо- и анацидные состояния ограничивают гидролиз пищевых белков, что затрудняет их дальнейшее переваривание. Наиболее выражено нарушение переваривания белков после обширной резекции желудка. Недостаточное расщепление пищевых белков наблюдается также при выпадении действия ферментов сока поджелудочной железы вследствие закупорки или сдавления ее выводящего протока. При энтеритах и энтероколитах усвоение пищевых белков ограничивается из-за ослабления секреторной и ускорения моторной функции тонкой кишки, а также нарушения ее всасывательной способности. При недостаточном питании или питании преимущественно растительными белками и при нарушении усвоения пищевых белков ограничение синтеза разнообразных белков в организме наступает не только из-за количественного недостатка аминокислот, но и из-за нарушения соотношения в содержании отдельных незаменимых аминокислот (дисбаланс). При выраженной белковой недостаточности наступает состояние отрицательного азотистого баланса, при к-ром количество выделяющегося из организма азота больше, чем количество азота, поступающего в организм.
Причиной расстройства белкового обмена в форме усиленного распада является нарушение регуляции метаболизма белковых структур. Ослабление и выпадение нервных воздействий на ткани приводит к нарушению их трофики и развитию трофических язв. Недостаток гормонов анаболического действия (соматотропный гормон, инсулин, половые гормоны) сопровождается первичным ослаблением биосинтеза белков. Недостаток соматотропного гормона у детей вызывает выраженное торможение роста. Дефицит инсулина при некомпенсированном сахарном диабете приводит к преобладанию процессов распада и отрицательному азотистому балансу. Первичное усиление распада белков наблюдается при тиреотоксикозе, избыточном действии стероидных гормонов коры надпочечников.
Усиление распада белков в тканях происходит также при повреждениях тканей (травмы, воспаление, аллергическая альтерация, ишемия, дегенерация). При общей интоксикации, в частности инфекционного происхождения, и обширных травмах мягких тканей и трубчатых костей преобладание распада в метаболизме белковых структур имеет генерализованный характер. Известную роль в этом играют продукты распада, поступающие из поврежденных тканей в общую циркуляцию.
Патология белкового обмена, кроме нарушения соответствия процессов синтеза и распада, в отношении отдельных видов белков проявляется также в форме врожденной недостаточности их биосинтеза, вследствие чего развивается, например, агаммаглобулинемия (см.), анальбуминемия (см.). Патология обмена белков может быть также в форме извращенного синтеза отдельных видов белков, она проявляется в образовании аномальных по своей структуре белков - некоторые виды гемоглобинопатии, появление белка Бене-Джонса (см. Бенс-Джонса белок), парапротеинов при миеломной болезни (см.) и другое.
Патология обмена аминокислот. Патология трансаминирования в форме недостаточности этого процесса возникает при уменьшении активности ферментов - трансаминаз (см. Ферменты), осуществляющих перенос аминогруппы с аминокислоты на α-кетокислоту. Такое нарушение имеет место при алиментарном гипо- или авитаминозе B6, поскольку витамин B6 является предшественником фосфопиридоксаля, а последний представляет собой активную группу (кофермент) трансаминаз. Абсолютной алиментарной недостаточности витамина В6 практически не встречается. Относительная недостаточность его поступления в организм может развиваться при повышенной потребности в нем, например, при беременности или при значительном подавлении антибиотиками и сульфамидными препаратами нормальной микрофлоры кишечника, где синтезируется витамин В6 в количестве, лишь частично покрывающем суточную потребность организма.
Недостаточность фосфопиридоксаля в организме может развиваться также вследствие нарушения ферментных систем, превращающих витамин B6 в его активную форму (метаболический авитаминоз), что может наблюдаться, по-видимому, при лечении фтивазидом больных туберкулезом.
Уменьшение активности трансаминаз может возникнуть и вследствие нарушения синтеза белковых структур трансаминаз (при белковой недостаточности) или изменения их конфигурации (связывание функциональных групп циклосерином, используемым при лечении туберкулеза). Локальное нарушение трансаминирования в отдельных органах возникает при повреждении их клеточных структур, особенно при некрозе последних. Это сопровождается выходом в кровь внутриклеточных ферментов и повышением в крови активности отдельных трансаминаз, определение которых используется в клинике в диагностических целях. Нарушение трансаминпрования в самих поврежденных органах носит при этом комплексный характер, оно обусловлено не только потерей ферментов из клеток, но и нарушением их биосинтеза, в том числе и ферментов синтеза фосфопиридоксаля.
Изменение интенсивности процесса трансаминирования в организме происходит и в результате нарушения соотношения реагирующих субстратов. При недостатке α-кетокислот, что может иметь место при угнетении цикла Кребса (например, при гипоксии, диабете), трансаминирование угнетается, а при избытке аминокислот, наблюдающемся при усиленном распаде белков, трансаминирование может быть усиленным. В последнем случае может возникать вторичное угнетение окисления в цикле Кребса (см. Окисление биологическое , Трикарбоновых кислот цикл).
Фактором нарушения трансаминирования может явиться расстройство регуляции активности отдельных трансаминаз под влиянием гормонов щитовидной железы и коры надпочечников.
Угнетение процесса дезаминирования может наступать в связи с причинами, вызывающими ослабление процесса трансаминирования, так как многие аминокислоты быстрее теряют свою аминогруппу в реакции трансаминирования с α-кетоглутаровой кислотой, чем в реакции прямого окислительного дезаминирования. Глутаминовая кислота, образующаяся при аминировании α-кетоглутаровой кислоты, быстрее, чем все остальные аминокислоты, подвергается окислительному дезаминированию с образованием аммиака. Этому способствует наличие в клетках специфического фермента - глутаматдегидрогеназы, функционирующей с участием НАД. В ряде исследований показано, что при экспериментальном авитаминозе B 6 или при инактивации фосфопиридоксаля тубазидом содержание отдельных аминокислот, кроме глутаминовой, в крови увеличивается, а образование мочевины в печени уменьшается.
Угнетение окислительного дезаминирования в печени возникает также вследствие ослабления биосинтеза белковых структур соответствующих ферментов при белковой недостаточности.
Ослабление окислительного дезаминирования наблюдается также при различных формах гипоксии (геморрагический шок и другое).
Следствием нарушения дезаминирования являются гипераминоацидемия (см. Аминоацидемия) - увеличение доли азота аминокислот в составе остаточного азота (см. Азот остаточный) и даже общая гиперазотемия и аминоацидурия (см.).
Наиболее отчетливо эти сдвиги в азотистом обмене наблюдаются при обширных поражениях клеток печени, особенно при гипоксии органа, когда нарушается не только процесс дезаминирования аминокислот, но и процесс мочевинообразования. При этом в составе остаточного азота, содержание которого может значительно возрастать, увеличивается концентрация азота аминокислот, а относительное (или даже абсолютное) количество азота мочевины уменьшается (продукционная гиперазотемия).
Продукционная гиперазотемия возникает и при патологических состояниях, сопровождающихся массивным распадом белков в организме. В этих условиях дезаминирование аминокислот и мочевинообразование в печени могут быть относительно недостаточными, и содержание остаточного азота крови будет увеличиваться за счет свободных аминокислот.
Увеличение содержания остаточного азота происходит также и при нарушении выделительной функции почек. Однако в этих условиях гиперазотемия происходит главным образом за счет увеличения в крови концентрации мочевины (ретенционная гиперазотемия). Клинической формой выраженной ретенционной гиперазотемии является уремия (см.). Гиперазотемии могут иметь и смешанный генез при одновременной недостаточности функции почек и печени и усиленном распаде белков. Продукционная гиперазотемия, не осложненная нарушением выделительной функции почек, приводит к потере аминокислот организма с мочой, так как фильтрация аминокислот в клубочковом аппарате почек превышает в этих условиях возможности их реабсорбции в канальцах (см. Почки). Усиленное выведение аминокислот выявлено в условиях белкового голодания при раневом истощении, травматических повреждениях трубчатых костей, спинного и головного мозга, в тяжелых случаях ожога, при инфекционных заболеваниях, в стадии кахексии при злокачественных новообразованиях, при гипертиреозе, болезни Иценко-Кушинга, длительном лечении глюкокортикоидами и препаратами АКТГ. В этих случаях гипераминоацидурия отражает в основном относительную недостаточность процессов дезаминирования избыточно освобождающихся при распаде белка аминокислот. Не исключено, что при этих состояниях может иметь место и прямое угнетение процессов дезаминирования в отдельных тканях, особенно в печени.
Другая группа гипераминоацидурий объединяет различные по происхождению формы нарушения обмена аминокислот, при которых увеличение их выделения связано с нарушением реабсорбции в канальцевой системе почек.
Генерализованное нарушение реабсорбции аминокислот происходит при фильтрации их из крови не в свободном состоянии, а в комплексе с металлами. Показано, что аминокислоты крови легко образуют комплексы с медью, свинцом, кадмием, ураном и при этом выводятся из организма.
При болезни Вильсона - Коновалова или гепато-лентикулярной дегенерации (см. Гепато-церебральная дистрофия), для которой характерно нарушение обмена меди, наблюдается значительная экскреция аминокислот в комплексе с медью без одновременного увеличения концентрации аминоазота в крови.
Нарушение почечной реабсорбции аминокислот имеет место и при синдроме Фанкони (см. Цистиноз), называемом некоторыми авторами аминовым диабетом. Для этого заболевания характерно сочетание усиленного выведения аминокислот (количество аминоазота в моче увеличивается в 30-40 раз) с гиперфосфатурией и псевдорахитическими изменениями в костях. Наблюдается также почечная глюкозурия (см. Диабет почечный).
Избирательное нарушение реабсорбции известно в отношении цистина. Однако цистинурия (см.) обычно сопровождается общим нарушением обмена этой аминокислоты. Описана врожденная аномалия обмена цистина, проявляющаяся в резко выраженной цистинурии без повышения содержания цистина в крови. Выведение цистина с мочой достигает в этих случаях 400-800 мг в сутки, тогда как нормальная экскреция цистина не превышает 80 мг. Цистин сравнительно плохо растворим, и увеличение его экскреции сопровождается образованием цистиновых камней в мочевыводящих путях.
Более тяжелое нарушение обмена цистина известно под названием цистиноза (см.). Это заболевание сопровождается общей аминоацидурией, в том числе и цистинурией, отложением кристаллов цистина в элементах ретикулоэндотелиальной системы; при нем наблюдается ранний смертельный исход.
Угнетение превращения фенилаланина в тирозин относится к наследственным заболеваниям. В крови и моче значительно увеличивается количество фенилаланина и ряда промежуточных продуктов его обмена, в частности фенилпировиноградной и фенилуксусной кислот. Клинически это нарушение обмена проявляется значительным отставанием умственного развития - фенилпировиноградная олигофрения (см. Фенилкетонурия). Недостаточно полное превращение фенилпировиноградной и фенилуксусной кислот в фенилацетилглутамин - нормальный конечный продукт обмена той части фенилаланина, которая не подвергалась превращению в тирозин, - выявлено и при вирусном гепатите. Ограниченное образование фенилацетилглутамина в этих случаях обусловлено первичным ограничением образования глутамина в печени.
Нарушение окислительного превращения тирозина в конечные продукты его обмена (фумаровую и ацетоуксусную кислоты) может сопровождаться накоплением различных промежуточных продуктов. Так, нарушение первого этапа этого пути обмена (переаминирования с α-кетоглутаровой к-той) приводит к гипертирозинемии, тярозинурии и состоянию тирозиноза (см.). Этот механизм нарушения обмена выявлен при экспериментальной белковой недостаточности, поражении печени четыреххлористым углеродом и экспериментальном лейкозе у мышей. В клинике аналогичное нарушение обмена тирозина наблюдается у больных лейкозом и при коллагенозах. Другая форма нарушения обмена тирозина - алкаптонурия (см.), развивающаяся при задержке окислительного превращения тирозина на стадии гомогентизиновой кислоты (см.). Патология относится к врожденным аномалиям обмена.
Нарушение других направлений в обмене тирозина также связано с активацией или угнетением ферментов, катализирующих реакции его специфических превращений. Превращение тирозина через стадию ДОФА в пигменты (меланины), окрашивающие кожу и волосы, определяется активностью тирозиназы, представляющей собой специфический медьсодержащий белок. Активность тирозиназы регулируется меланофорным гормоном гипофиза, синтез которого сдерживается гормонами коры надпочечников. При гипофункции надпочечников могут возникать нарушения пигментного обмена (см.). Альбинизм (см.) представляет собой врожденную аномалию обмена тирозина, состоящую в выпадении синтеза фермента тирозиназы.
Основной путь обмена триптофана в организме заканчивается превращением его в никотиновую кислоту. Ряд промежуточных продуктов на этом пути обмена триптофана, а именно 3-оксикинуренин, ксантуреновая, 3-оксиантраниловая кислоты и их производные, обладают при повышенной концентрации патогенными свойствами.
Ксантуреновая кислота способствует распаду гликогена и гипергликемии. При длительном повышении ее концентрации в крови у экспериментальных животных наблюдаются дегенеративные изменения в бетаклетках поджелудочной железы. 3-Оксикинуренин и 3-оксиантраниловая кислоты могут проявлять также канцерогенное действие.
Накопление в крови промежуточных продуктов обмена триптофана происходит вследствие подавления активности ряда ферментов, функционирующих в комплексе с производными витаминов В6, B1, В2 и PP. Избыточное образование токсических метаболитов выявлено при хроническом гепатите, тяжелых формах сахарного диабета, остром лейкозе, хронических миело- и лимфолейкозе, лимфогранулематозе, ревматизме и склеродермии. Нарушение обмена триптофана может быть выявлено при помощи пробы с нагрузкой.
Увеличение концентрации креатинина в крови происходит при нарушении выделения его почками, а увеличенное пли уменьшенное выведение его с мочой, без одновременной ретенции в крови, отражает нарушение образования его из креатина при патологии обмена последнего в мышечной ткани. Увеличение экскреции креатинина наблюдается при гипофункции щитовидной железы. Уменьшение экскреции креатинина в сочетании с увеличенным выведением креатина имеет место при гипертиреозе, тяжелом течении сахарного диабета и особенно при миопатиях (миастения, миозит, миотония).
Количество азота мочевой кислоты - конечного продукта пуринового обмена (см.) - в составе остаточного азота крови колеблется от 0,1 до 3.0 мг%. Патологическое увеличение его концентрации наблюдается при массивном распаде клеточных структур (голодание, тяжелая мышечная работа, инфекции и так далее), когда усиление эритропоэза (см. Кроветворение) сопровождается освобождением ядер из ретикулоцитов. Выведение мочевой кислоты с мочой ограничивается ее интенсивной реабсорбцией. Увеличение концентрации мочевой кислоты в крови обусловливает возможность отложения ее в хрящах, суставных сумках, сухожилиях, фасциях, а иногда в почках, мышцах и коже.
Азотистый обмен в облученном организме
Характер изменений азотистого обмена в основном зависит от дозы облучения. При воздействии больших доз ионизирующей радиации в организме происходит процесс патологического распада белков органов и тканей, которые не восстанавливаются белками пищи, что проявляется в отрицательном азотистом балансе, особенно при облучении в летальных дозах.
В изменении азотистого обмена в облученном организме существенную роль играет пониженное всасывание аминокислот стенками тонкой кишки, а также повышенное выделение азота с мочой в ближайшие дни после лучевого поражения. Так, например, при тотальном воздействии гамма-излучением и нейтронами выделение аминокислот с мочой у людей увеличивается в 10 раз по сравнению с нормой. При облучении в больших дозах экспериментальных животных отмечалось увеличение содержания мочевины в моче, тирозина в крови, выведение аминокислот с мочой, креатинурия (см.), что указывает на усиление тканевого распада. Увеличение распада белков может быть также результатом повышения активности протеолитических ферментов. В свою очередь повышение активности протеолитических ферментов связывают с непосредственным поражением внутриклеточных мембран.
Изменения азотистого обмена при облучении зависят от следующих основных причин: непосредственное воздействие радиации на молекулы белка в клетке и изменение его физико-химических свойств; изменение биохимических механизмов синтеза белка; интенсификация протеолитических ферментов в клетке; опосредованное влияние радиации на деятельность желез внутренней секреции и так далее
В пожилом возрасте существенно снижается функциональная способность пищеварительного тракта (ослабляется синтез и секреция соляной к-ты, протеолитических ферментов), замедляется всасывание свободных аминокислот в кишечнике; снижается способность ассимиляции пищевых веществ на тканевом и клеточном уровнях, что связано в первую очередь с дизадаптацией ферментных систем организма; нарушаются процессы биосинтеза белков, нуклеиновых кислот и так далее.
По мере старения организма снижается его способность ассимилировать белки, увеличиваются эндогенные потери белковых компонентов пищи, что характеризуется появлением отрицательного азотистого баланса.
Причины снижения интенсивности синтеза белка в старости до сих пор остаются неясными. Большинство исследователей считает, что при старении первичные изменения возникают в регуляторных генах, приводя в одних случаях к нарастающему подавлению транскрипции отдельных оперонов (см.), а в других - к временному усилению биосинтеза некоторых белков. При этом неравномерно изменяется биосинтез различных белков, сокращается возможный диапазон активации биосинтеза, быстрее нарастает снижение потенциальных возможностей биосинтетических систем в условиях напряженной деятельности. В последующем наступают изменения и в структурных генах, что приводит к определенным качественным сдвигам в синтезируемых белковых молекулах, в частности к изменениям в аллостерическом регулировании активности ферментов.
Характерным примером изменения азотистого обмена при старении является нарушение пуринового обмена, когда в крови и тканях накапливается большое количество уратов, которые отлагаются затем в суставах и хрящах (см. Подагра). Однако так наз. отложения солей связаны не только с нарушениями пуринового и минерального обмена.
Имеются убедительные доказательства, что причины отложения солей в суставах и хрящах заключаются не только в увеличении концентрации уратов и кальция, а прежде всего в изменении свойств структурных белков соединительной ткани, в частности коллагена. Качественные изменения белков проявляются главным образом в нарушении третичной и четвертичной структуры молекулы белка (см.). При этом отмечено повышение прочности белковой структуры, обусловленное появлением дополнительных, перекрестных связей между отдельными компонентами. В процессе старения меняются физико-химических свойства белков, в частности снижается лабильность, дисперсность, гидрофильность и электрический заряд их молекул. Гипотеза отечественных авторов (А. А. Богомолец, А. В. Нагорный, В. Н. Никитин) о важности для процессов старения изменения физико-химических свойств белков, например, белков соединительной ткани, их огрубения и понижения функциональной активности, находит все большее признание в мировой литературе.
Многочисленные экспериментальные и клинические наблюдения особенностей разбалансирования процессов азотистого обмена при старении послужили основанием для разработки специальных рационов. В основе рекомендаций для составления таких рационов, направленных на нормализацию нарушений азотистого обмена у лиц пожилого и старческого возраста, лежат: принцип энергетической сбалансированности пищевого рациона с энерготратами организма; обеспечение в рационах сравнительно высоких количеств белка (1,2-1,3 г на 1 кг веса) с высоким содержанием полноценных животных белков (главным образом белков молока); ограничение в рационах продуктов высокой концентрации пуриновых оснований (см.); обеспечение в рационах достаточного содержания витаминов и микроэлементов, в частности аскорбиновой к-ты, ниацина, тиамина, рибофлавина, кобаламина и другое, что необходимо для обновления изнашивающихся в процессе жизнедеятельности ферментных систем.
Особенности азотистого обмена у детей
Интенсивность процессов азотистого обмена на протяжении роста ребенка подвергается изменениям, особенно ярко выраженным у новорожденных и детей раннего возраста. В течение первых трех дней жизни баланс азота является отрицательным, что объясняется недостаточным поступлением белка с малым количеством пищи. В этот период обнаруживается транзиторное повышение остаточного азота в крови до 55-60 мг% . Количество выделяемого почками азота нарастает в течение первых 3 дней, после чего падает и начинает вновь увеличиваться со второй недели жизни параллельно возрастающему количеству пищи.
Общая особенность азотистого обмена у детей - положительный баланс азота, что является необходимым условием роста. Азот пищи в максимальной степени используется растущим организмом для пластических целей. Так, например, на ранних этапах развития детского организма ферментные системы, обеспечивающие синтез нуклеиновых кислот, отличаются наивысшей активностью, в то же время активность ферментов, катализирующих их распад, снижена.
Наиболее высокая усвояемость азота в организме наблюдается у детей первых месяцев жизни. Баланс азота заметно снижается в период 3-6 месяцев жизни, хотя и остается положительным.
Во втором полугодии жизни баланс азота стабилизируется. По данным В. Ф. Ведрашко (1958), у детей 2-3 лет, получающих 4-4,2 г/кг белка, баланс азота составляет 2,3 г, ретенция (т. е. задержка) - 30% при соотношении животных и растительных белков 4:1. У детей 4-6 лет удовлетворительный баланс и ретенция азота достигаются при получении 3,5 г/кг белка: баланс 2,7 г, ретенция - 25% (В. Ф. Ведрашко и Э. И. Аршавская, 1965). У детей 7-8 лет азотистое равновесие достигается при введении 2,5 г/кг белка: баланс 2,8-3 г, ретенция - в пределах 21% . По данным Ин-та питания АМН СССР, у детей 11-13 лет при введении 2 г/кг белка азотистый баланс составляет 1,8 г, ретенция - 13,8%.
Показатели ретенции и баланса азота подвержены значительным индивидуальным колебаниям, зависят от количества белка пищи, его соотношения с другими пищевыми ингредиентами. Установлены также сезонные колебания этих показателей: они выше в весеннее и летнее время и ниже зимой.
Потребность в незаменимых аминокислотах у детей выше, чем у взрослых, при этом для детского организма к незаменимым аминокислотам относят и гистидин. Средние величины потребности в незаменимых аминокислотах, по данным ФАО ВОЗ(1963), представлены в таблице 3.
Таблица 3. Потребности в незаменимых аминокислотах у детей
Клетки растущих тканей отличаются высокой концентрацией аминокислот, что свидетельствует о высокой активности механизмов, обеспечивающих транспорт аминокислот через клеточные мембраны. Активный мембранный транспорт аминокислот имеет место в плаценте. Дент (С. Е. Dent, 1948) говорит в связи с этим о «плацентарном аминокислотном насосе», обеспечивающем движение аминокислот от матери к плоду (см. Плацента). Показано, что этот процесс отличается строгой стереоспецифичностью, то есть левовращающие (L-аминокислоты) проходят плацентарный барьер с более высокой скоростью, чем правовращающие (D-аминокислоты). Функция плаценты позволяет объяснить более высокое содержание аминокислот в пуповинной крови по сравнению с кровью детей более старшего возраста и взрослых (табл. 4).
Таблица 4. Содержание свободных аминокислот в крови (в мг%) (по Шрейеру, 1965)
| Аминокислота | В пуповинной крови | В крови детей | В крови взрослых |
|---|---|---|---|
| Аланин | 4,8 | 3,9 | 3,8 |
| Аргинин | 3,3 | 2,2 | 2,1 |
| Глицин | 3,4 | 2,6 | 2,8 |
| Гистидин | 3,4 | 1,8 | 1,7 |
| Изолейцин | 2,3 | 1,7 | 1,6 |
| Лейцин | 2,5 | 2,3 | 2,0 |
| Лизин | 8,1 | 2,4 | 2,8 |
| Метионин | 0,5 | 0,3 | 0,35 |
| Фенилаланин | 2,3 | 1,6 | 1,6 |
| Треонин | 2,8 | 2,3 | 2,0 |
| Триптофан | 1,7 | 0,8 | 1,1 |
| Тирозин | 2,3 | 1,6 | 1,4 |
| Валин | 4,9 | 3,2 | 3,0 |
Существенное влияние на рост ребенка оказывают дефекты питания, вследствие которых ребенок получает избытки отдельных аминокислот, что служит причиной задержки физического развития, гипераминоацидурии, интоксикации.
У детей раннего возраста повышена экскреция аминокислот с мочой - так называемая физиологическая гипераминоацидурия. На первой неделе жизни азот аминокислот составляет 3-4% от общего азота мочи (по некоторым данным, до 10%) и лишь к концу первого года жизни снижается до 1 %. В этот период выведение аминокислот в расчете на 1 кг веса достигает величин выведения их у взрослого человека, экскреция аминоазота, достигающая у новорожденных 10 мг/кг, на втором году жизни редко превышает 2 мг/кг. В моче новорожденных повышено по сравнению со взрослыми содержание таурина, треонина, серина, глицина, аланина, цистина, лейцина, тирозина, фенилаланина и лизина. В первые месяцы жизни в моче обнаруживается также этаноламин и гомоцитруллин. В моче детей первого года жизни преобладают аминокислоты пролин и гидроксипролин. Причиной физиологической гипераминоацидурии является функциональная незрелость почек, проявляющаяся в недостаточной реабсорбции аминокислот из клубочкового фильтрата (гипераминоацидурия ренального типа). Доказательством этого служит более высокий клиренс аминокислот. У недоношенных, кроме того, имеет место гипераминоацидурия перегрузочного типа, так как содержание свободных аминокислот в плазме крови выше, чем у доношенных.
В процессе роста ребенка меняются количественные и качественные характеристики азотистого обмена
Еще Вирхов (R. Virchow, 1856) обратил внимание на то, что моча плода содержит избытки мочевой кислоты и лишь следы мочевины. Отложение кристаллов мочевой кислоты в почечной ткани он назвал мочекислым инфарктом новорожденных. Исследования важнейших азотистых компонентов мочи у детей показали, что соотношение мочевой кислоты, мочевины и аммиака в процессе роста существенно изменяется. Так, первые 3 месяца жизни характеризуются наименьшим содержанием в моче мочевины и наибольшей экскрецией мочевой кислоты. В возрасте от 3 до 6 месяцев в моче нарастает количество мочевины и снижается содержание мочевой кислоты. Выведение мочевой кислоты на протяжении первого - второго года жизни в расчете на 1 кг веса превышает таковое у взрослых, содержание аммиака в моче в первые дни жизни невелико, но затем резко возрастает и держится на высоком уровне на протяжении всего первого года жизни. Н. Ф. Толкачевская (1960) связывает эти особенности азотистого обмена с преобладанием у плода и новорожденного урикотелического пути обмена аммиака (нейтрализация аммиака обеспечивается главным образом за счет усиленного образования мочевой кислоты). Это филогенетически более древний путь, который на первом году жизни постепенно и почти полностью вытесняется уреотелическим - синтезом мочевины в цикле Кребса-Гензелейта.
Важной особенностью азотистого обмена у детей является физиологическая креатинурия (см.). Креатин обнаруживается в амниотической жидкости и в моче, начиная с периода новорожденности и до периода полового созревания. Суточная экскреция креатинина с возрастом увеличивается, в то же время по мере нарастания массы тела азот креатинина в процентном отношении к общему азоту мочи снижается. Количество выведенного с мочой креатинина в расчете на 1 кг веса у детей колеблется в пределах 5,5-10 мг. Величины суточного выведения креатинина, процентного отношения к общему азоту мочи, полученные у разных детей одного возраста, близки между собой.
Библиография: Белозерский А. Н. Молекулярная биология - новая ступень познания природы, М., 1970; Браунштейн А. Е. Биохимия аминокислотного обмена, М., 1949, библиогр.; Збарский Б. И., Иванов И. И. и Мардашев С. Р. Биологическая химия, Л., 1972; Иванов И. И. и др. Введение в клиническую биохимию, Л., 1969, библиогр.; Химия и биохимия нуклеиновых кислот, под ред. И. Б. Збарского и С. С. Дебова, Л., 1968; Lehninger A. L. Biochemistry, N. Y., 1970.
Радиоизотопное исследование азотистого обмена - Белки, под ред. Г. Нейрата и К. Бэйли, пер. с англ., т. 3, ч. 2, с. 594, М., 1950, библиогр.; Хаггис Д ж. и др. Введение в молекулярную биологию, пер. с англ., с. 341, М., 1967.
Патология азотистого обмена . - Ангелов А. М. и др. Влияние тироксина на активность некоторых ферментов обмена углеводов и аминокислот в печени морских свинок, Вопр. мед. хим., т. 17, в. 2, с. 165, 1971, библиогр.; Бадалян Л. О., Таболин В. А. и Вельтищев Ю. Е. Наследственные болезни у детей, с. 44, М., 1971; Капланский С. Я. Вопросы патологии обмена белков и аминокислот, в кн.: Хим. основы процессов жизнедеятельности, под ред. В. Н. Ореховича, с. 253, М., 1962, библиогр.; он же, Патологическая физиология белкового обмена, Многотомн. руководство по пат. физиол., под ред. Н. Н. Сиротинина, т. 2, с. 455, М., 1966, библиогр.; Кремер Ю. Н. Биохимия белкового питания, Рига, 1965, библиогр.; Лаптева Н. Н. Патофизиология белкового обмена, М., 1970, библиогр.; Майстер А. Биохимия аминокислот, пер. с англ., с. 463, М., 1961; Мардашев С. Р. и д р. Опыт и перспективы лечения L-глютамином больных тяжелым эпидемическим гепатитом (болезнь Боткина), в кн.: Усп. гепатол., под ред. Е. М. Тареева и А. Ф. Блюгера, с. 401, Рига, 1971; Репин И. С. Азотистый обмен при лихорадочных состояниях, Л., 1961, библиогр.; Xорст А. Молекулярная патология, пер. с польск., с. 196, М., 1967, библиогр.
Азотистого обмена в облученном организме - Бак З. и Александер П. Основы радиобиологии, пер. с англ., М., 1963, библиогр.; Ранние радиационно-биохимические реакции, под ред. Е. Ф. Романцева, М., 1966, библиогр.; Штреффер К. Радиационная биохимия, пер. с нем., М., 1972.
Изменение азотистого обмена в процессе старения - Ведущие проблемы советской геронтологии, под ред. Д. Ф. Чеботарева и др., Киев, 1972; Ведущие проблемы возрастной физиологии и биохимии, под ред. В. Н. Никитина, М., 1966, библиогр.; Парина Е. В. Возраст и обмен белков, Харьков, 1967, библиогр.; Фролькис В. В. Регулирование, приспособление и старение, Л., 1970, библиогр.; Gsеll D. Protein and nitrogen metabolism in the old age, Proc. 4-th int. congr. dietetics, p. 88, Stockholm, 1965; Verz#225;r F. Aging of the collagen fiber, Int. Rev. Connect. Tissue Res., v. 2, p. 243, 1964.
Азотистый обмен у детей - Питание здорового И больного ребенка, под ред. М. И. Олевского и Ю. К. Полтевой, с. 59, 212, М., 1965; Толкачевская Н. Ф. Развитие процессов обмена у детей первого года жизни, М., 1960, библиогр.; Тур А. Ф. Физиология и патология новорожденных детей, Л., 1967; Schreier K. Eiweiβstoffwechsel, Handb. d. Kinderheilk., hrsg. v. H. Opitz u. P. Schmid, S. 57, B. u. a., 1965, Bibliogr.; Sсhreiеr К. u. a. Über die Clearance-Rate einiger Aminosäuren bei Säuglingen und Frühgeborenen, Z. Kinderheilk., Bd 79, S. 165, 1957, Bibliogr.; SereniF. a. Principi N. The development of enzyme systems, Pediat. Clin. N. Amer., v. 12, p. 515, 1965, bibliogr.
С. Е. Северин, Н. Н. Лаптева; Ю. Е. Вельтищев (пед.), Г. И. Козинец (рад.), Е. Ф. Романцев (рад. био.), В. А. Тутельян (гер.).
 Азотистый обмен - совокупность химических превращений азотсодержащих веществ в организме. А. о. включает обмен простых и сложных белков, нуклеиновых кислот, продуктов их распада (пептидов, аминокислот и нуклеотидов), содержащих азот жироподобных веществ (липидов), аминосахаров, гормонов, витаминов и др. Для нормального течения процессов жизнедеятельности организм должен быть обеспечен необходимым количеством усвояемого азота. Главнейшей составной частью и основным источником азота пищи человека являются белковые вещества
Азотистый обмен - совокупность химических превращений азотсодержащих веществ в организме. А. о. включает обмен простых и сложных белков, нуклеиновых кислот, продуктов их распада (пептидов, аминокислот и нуклеотидов), содержащих азот жироподобных веществ (липидов), аминосахаров, гормонов, витаминов и др. Для нормального течения процессов жизнедеятельности организм должен быть обеспечен необходимым количеством усвояемого азота. Главнейшей составной частью и основным источником азота пищи человека являются белковые вещества

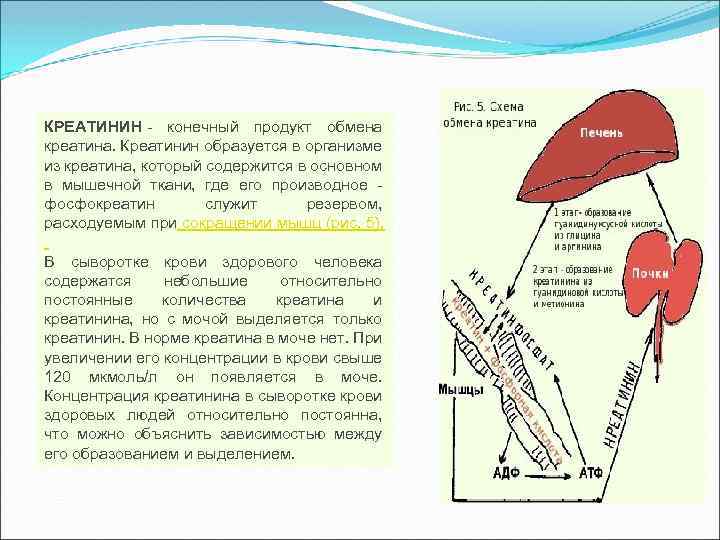 КРЕАТИНИН - конечный продукт обмена креатина. Креатинин образуется в организме из креатина, который содержится в основном в мышечной ткани, где его производное - фосфокреатин служит резервом, расходуемым при сокращении мышц (рис. 5). В сыворотке крови здорового человека содержатся небольшие относительно постоянные количества креатина и креатинина, но с мочой выделяется только креатинин. В норме креатина в моче нет. При увеличении его концентрации в крови свыше 120 мкмоль/л он появляется в моче. Концентрация креатинина в сыворотке крови здоровых людей относительно постоянна, что можно объяснить зависимостью между его образованием и выделением.
КРЕАТИНИН - конечный продукт обмена креатина. Креатинин образуется в организме из креатина, который содержится в основном в мышечной ткани, где его производное - фосфокреатин служит резервом, расходуемым при сокращении мышц (рис. 5). В сыворотке крови здорового человека содержатся небольшие относительно постоянные количества креатина и креатинина, но с мочой выделяется только креатинин. В норме креатина в моче нет. При увеличении его концентрации в крови свыше 120 мкмоль/л он появляется в моче. Концентрация креатинина в сыворотке крови здоровых людей относительно постоянна, что можно объяснить зависимостью между его образованием и выделением.
 Мочевина – конечный продукт распада белка в организме, при выведении которого через почки осуществляется удаление остатков «ненужного» азота. Образуется в печени. Выводится вместе с мочой и частично с потом (что придает поту специфический запах). Мочевина помогает организму сохранять воду и некоторые микроэлементы. Это выполняется посредством повторного всасывания почками в кровоток. Каждый раз, когда происходит процесс всасывания, мочевина «тянет» за собой обратно в кровь молекулы воды и полезных минералов. Однако избыточное ее содержание наносит вред органам и тканям.
Мочевина – конечный продукт распада белка в организме, при выведении которого через почки осуществляется удаление остатков «ненужного» азота. Образуется в печени. Выводится вместе с мочой и частично с потом (что придает поту специфический запах). Мочевина помогает организму сохранять воду и некоторые микроэлементы. Это выполняется посредством повторного всасывания почками в кровоток. Каждый раз, когда происходит процесс всасывания, мочевина «тянет» за собой обратно в кровь молекулы воды и полезных минералов. Однако избыточное ее содержание наносит вред органам и тканям.
 Мочевая кислота Конечный продукт метаболизма пуриновых оснований, входящих в состав нуклеотидов. Благодаря выведению мочевой кислоты из организма удаляется избыток азота. В плазме крови мочевая кислота содержится преимущественно в форме натриевой соли. Концентрация мочевой кислоты в крови обусловлена равновесием процессов синтеза мочевой кислоты и её выведения почками.
Мочевая кислота Конечный продукт метаболизма пуриновых оснований, входящих в состав нуклеотидов. Благодаря выведению мочевой кислоты из организма удаляется избыток азота. В плазме крови мочевая кислота содержится преимущественно в форме натриевой соли. Концентрация мочевой кислоты в крови обусловлена равновесием процессов синтеза мочевой кислоты и её выведения почками.
 ПУТИ ОБРАЗОВАНИЯ АММИАКА глутаминовая кислота -кетоглутаровая кислота + NH 3 -аминокислота -кетокислота + NH 3 цистеин пируват + NH 3 гистидин урокаиновая кислота + NH 3 глицин глиоксалевая кислота + NH 3 глюкозамин-6 -фосфат глюкоза-6 -фосфат + NH 3 глутамин глутаминовая кислота + NH 3 O ║ NH 2 -C O-P +CO 2 + NH 3 карбомоилфосфат глутамин АТФ АДФ мочевина пиримидины фолиевая кислота пурины глюкозамин
ПУТИ ОБРАЗОВАНИЯ АММИАКА глутаминовая кислота -кетоглутаровая кислота + NH 3 -аминокислота -кетокислота + NH 3 цистеин пируват + NH 3 гистидин урокаиновая кислота + NH 3 глицин глиоксалевая кислота + NH 3 глюкозамин-6 -фосфат глюкоза-6 -фосфат + NH 3 глутамин глутаминовая кислота + NH 3 O ║ NH 2 -C O-P +CO 2 + NH 3 карбомоилфосфат глутамин АТФ АДФ мочевина пиримидины фолиевая кислота пурины глюкозамин
Src="http://present5.com/presentation/229073585_437016682/image-8.jpg" alt="ПРИЧИНЫ АММИАЧНОЙ ИНТОКСИКАЦИИ ГИПЕРАММОНИЙЕМИЯ (>25 -40 мкмоль/л) Нарушение мочевинообразующей функции печени Нарушение азотовыделительной функции"> ПРИЧИНЫ АММИАЧНОЙ ИНТОКСИКАЦИИ ГИПЕРАММОНИЙЕМИЯ (>25 -40 мкмоль/л) Нарушение мочевинообразующей функции печени Нарушение азотовыделительной функции почек Острая почечная недостаточность Вирусный гепатит Хроническая почечная недостаточность Токсический гепатит Цирроз печени Портакавальные анастомозы на фоне богатой белками пищи Врожденные гипераммонийемии
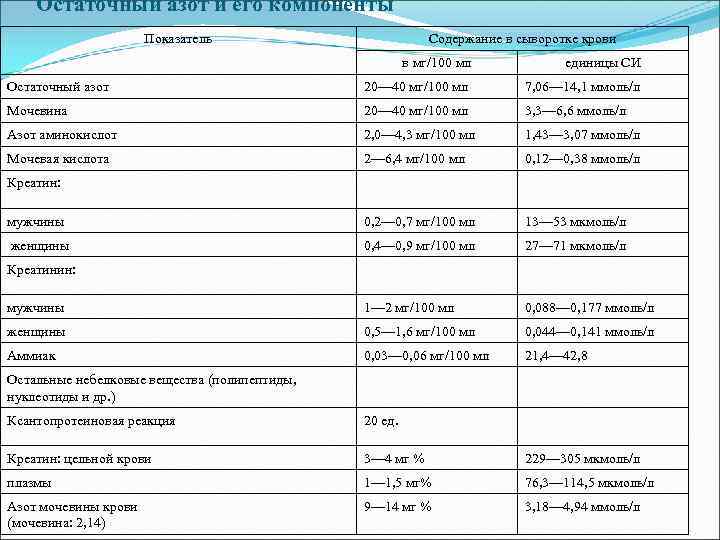 Остаточный азот и его компоненты Показатель Содержание в сыворотке крови в мг/100 мл единицы СИ Остаточный азот 20- 40 мг/100 мл 7, 06- 14, 1 ммоль/л Мочевина 20- 40 мг/100 мл 3, 3- 6, 6 ммоль/л Азот аминокислот 2, 0- 4, 3 мг/100 мл 1, 43- 3, 07 ммоль/л Мочевая кислота 2- 6, 4 мг/100 мл 0, 12- 0, 38 ммоль/л мужчины 0, 2- 0, 7 мг/100 мл 13- 53 мкмоль/л женщины 0, 4- 0, 9 мг/100 мл 27- 71 мкмоль/л мужчины 1- 2 мг/100 мл 0, 088- 0, 177 ммоль/л женщины 0, 5- 1, 6 мг/100 мл 0, 044- 0, 141 ммоль/л Аммиак 0, 03- 0, 06 мг/100 мл 21, 4- 42, 8 Креатин: Креатинин: Остальные небелковые вещества (полипептиды, нуклеотиды и др.) Ксантопротеиновая реакция 20 ед. Креатин: цельной крови 3- 4 мг % 229- 305 мкмоль/л плазмы 1- 1, 5 мг% 76, 3- 114, 5 мкмоль/л Азот мочевины крови (мочевина: 2, 14) 9- 14 мг % 3, 18- 4, 94 ммоль/л
Остаточный азот и его компоненты Показатель Содержание в сыворотке крови в мг/100 мл единицы СИ Остаточный азот 20- 40 мг/100 мл 7, 06- 14, 1 ммоль/л Мочевина 20- 40 мг/100 мл 3, 3- 6, 6 ммоль/л Азот аминокислот 2, 0- 4, 3 мг/100 мл 1, 43- 3, 07 ммоль/л Мочевая кислота 2- 6, 4 мг/100 мл 0, 12- 0, 38 ммоль/л мужчины 0, 2- 0, 7 мг/100 мл 13- 53 мкмоль/л женщины 0, 4- 0, 9 мг/100 мл 27- 71 мкмоль/л мужчины 1- 2 мг/100 мл 0, 088- 0, 177 ммоль/л женщины 0, 5- 1, 6 мг/100 мл 0, 044- 0, 141 ммоль/л Аммиак 0, 03- 0, 06 мг/100 мл 21, 4- 42, 8 Креатин: Креатинин: Остальные небелковые вещества (полипептиды, нуклеотиды и др.) Ксантопротеиновая реакция 20 ед. Креатин: цельной крови 3- 4 мг % 229- 305 мкмоль/л плазмы 1- 1, 5 мг% 76, 3- 114, 5 мкмоль/л Азот мочевины крови (мочевина: 2, 14) 9- 14 мг % 3, 18- 4, 94 ммоль/л
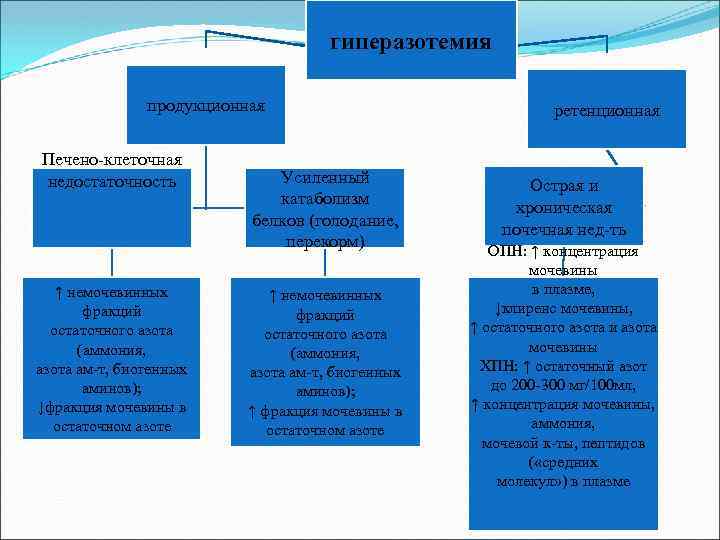 гиперазотемия продукционная Печено-клеточная недостаточность немочевинных фракций остаточного азота (аммония, азота ам-т, биогенных аминов); ↓фракция мочевины в остаточном азоте Усиленный катаболизм белков (голодание, перекорм) немочевинных фракций остаточного азота (аммония, азота ам-т, биогенных аминов); фракция мочевины в остаточном азоте ретенционная Острая и хроническая почечная нед-ть ОПН: концентрация мочевины в плазме, ↓клиренс мочевины, остаточного азота и азота мочевины ХПН: остаточный азот до 200 -300 мг/100 мл, концентрация мочевины, аммония, мочевой к-ты, пептидов («средних молекул») в плазме
гиперазотемия продукционная Печено-клеточная недостаточность немочевинных фракций остаточного азота (аммония, азота ам-т, биогенных аминов); ↓фракция мочевины в остаточном азоте Усиленный катаболизм белков (голодание, перекорм) немочевинных фракций остаточного азота (аммония, азота ам-т, биогенных аминов); фракция мочевины в остаточном азоте ретенционная Острая и хроническая почечная нед-ть ОПН: концентрация мочевины в плазме, ↓клиренс мочевины, остаточного азота и азота мочевины ХПН: остаточный азот до 200 -300 мг/100 мл, концентрация мочевины, аммония, мочевой к-ты, пептидов («средних молекул») в плазме
 ИСТОЧНИКИ И СПОСОБЫ ОБЕЗВРЕЖИВАНИЯ АММИАКА В РАЗНЫХ ТКАНЯХ Биогенные амины Аминокислоты Нуклеотиды АММИАК Синтез Образование мочевины глутамина аланина глутамата аммонийных (25 г/сут) солей (0, 5 г/сут) печень мозг Мышцы, кишечник мозг почки
ИСТОЧНИКИ И СПОСОБЫ ОБЕЗВРЕЖИВАНИЯ АММИАКА В РАЗНЫХ ТКАНЯХ Биогенные амины Аминокислоты Нуклеотиды АММИАК Синтез Образование мочевины глутамина аланина глутамата аммонийных (25 г/сут) солей (0, 5 г/сут) печень мозг Мышцы, кишечник мозг почки
 РЕАКЦИИ СВЯЗЫВАНИЯ АММИАКА В КЛЕТКЕ 1. Реакция восстановительного аминирования -кетоглутарата в L-глутамат: НАДФН 2 НАДФН NH 3 + -кетоглутаровая глутаминовая + Н 2 О кислота глутаматдегидрогеназа кислота 2. Реакция образования глутамина из глутаминовой кислоты с участием фермента глутаминсинтетазы. Реакция протекает в цитозоле клеток всех тканей, но в большей степени – мозга: COOH CONH 2 │ │ CH 2 АТФ АДФ + Фн CH 2 │ │ CH 2 2 CH │ │ HC-NH 2 глутаминсинтетаза HC-NH 2 │ │ COOH COOH глутаминовая глутамин кислота
РЕАКЦИИ СВЯЗЫВАНИЯ АММИАКА В КЛЕТКЕ 1. Реакция восстановительного аминирования -кетоглутарата в L-глутамат: НАДФН 2 НАДФН NH 3 + -кетоглутаровая глутаминовая + Н 2 О кислота глутаматдегидрогеназа кислота 2. Реакция образования глутамина из глутаминовой кислоты с участием фермента глутаминсинтетазы. Реакция протекает в цитозоле клеток всех тканей, но в большей степени – мозга: COOH CONH 2 │ │ CH 2 АТФ АДФ + Фн CH 2 │ │ CH 2 2 CH │ │ HC-NH 2 глутаминсинтетаза HC-NH 2 │ │ COOH COOH глутаминовая глутамин кислота
 Пути обмена азота аминокислот и аммиака ТКАНИ КРОВЬ ПЕЧЕНЬ Мышцы, кишечник Аминок-ты -КГ Ала -КГ NH 3 Кеток-ты глутамат пируват пируват глутамат СО 2 АМФ NH 3 глюкоза карбомоил фосфат ИМФ глутамат глутамин МОЗГ и другие ткани Аминок-ты -КГ NH 3 Кеток-ты глутамат глутамин -КГ ПОЧКИ NH 3 глутамат NH 3 МОЧА орнитиновый цикл NH 3 глутамат мочевина аммонийные соли глутамат мочевина
Пути обмена азота аминокислот и аммиака ТКАНИ КРОВЬ ПЕЧЕНЬ Мышцы, кишечник Аминок-ты -КГ Ала -КГ NH 3 Кеток-ты глутамат пируват пируват глутамат СО 2 АМФ NH 3 глюкоза карбомоил фосфат ИМФ глутамат глутамин МОЗГ и другие ткани Аминок-ты -КГ NH 3 Кеток-ты глутамат глутамин -КГ ПОЧКИ NH 3 глутамат NH 3 МОЧА орнитиновый цикл NH 3 глутамат мочевина аммонийные соли глутамат мочевина
 ЦИКЛ МОЧЕВИНЫ И ЕГО СВЯЗЬ С ЦТК цитоплазма NH 3 + CO 2 + Н 2 О L-аспартат 2 АТФ карбомоил-фосфат цитруллин аргининосукцинат оксалоацетат митохондрия малат митохондрия орнитин аргинин фумарат мочевина ЦТК
ЦИКЛ МОЧЕВИНЫ И ЕГО СВЯЗЬ С ЦТК цитоплазма NH 3 + CO 2 + Н 2 О L-аспартат 2 АТФ карбомоил-фосфат цитруллин аргининосукцинат оксалоацетат митохондрия малат митохондрия орнитин аргинин фумарат мочевина ЦТК
Форма выведения белкового азота - в виде аммиака, мочевины или же мочевой кислоты - тесно связана с условиями жизни цветного и наличием воды (табл. 10.4). Аммиак весьма токсичен даже в очень малых концентрациях, поэтому он должен быстро
удаляться либо путем выведения во внешнюю среду, либо путем превращения в менее токсичные вещества (мочевину или мочевую кислоту).
У большинства водных беспозвоночных конечным продуктом белкового обмена является аммиак. Благодаря его легкой растворимости и небольшому молекулярному весу он диффундирует чрезвычайно быстро. Значительная его часть может быть выделена через любую поверхность, соприкасающуюся с водой, - не обязательно через почку. У костистых рыб большая часть азота выводится в форме аммиака через жабры. У карпа и золотой рыбки жабры выделяют в 6-10 раз больше азота, чем почки, и только 10% его составляет мочевина; остальные 90% выводятся в виде аммиака (Smith, 1929).
МОЧЕВИНА
Мочевина легко растворима в воде и обладает довольно малой токсичностью. Синтез мочевины у высших животных был изучен знаменитым биохимиком Гансом Кребсом - тем самым ученым, по имени которого был назван цикл окислительного энергетического обмена (цикл трикарбоновых кислот, или цикл Кребса),
При синтезе мочевины аммиак и двуокись углерода, конденсируясь с фосфатом, образуют карбамоилфосфат, который затем используется для синтеза цитруллина из орнитина, как показано на рис. 10.13. После этого добавляется еще одна молекула аммиака из аспарагиновой кислоты, и это ведет к образованию аминокислоты аргинина. В присутствии фермента аргиназы аргинин распадается на мочевину и орнитин. Из орнитина синтезируется новая молекула цитруллина, и весь цикл повторяется; поэтому весь этот путь превращений называют орнитиновым циклом синтеза мочевины. Наличие аргиназы у животного говорит о его способности вырабатывать мочевину и часто указывает на то, что мочевина является у него главным азотистым экскретом. Но это необязательно так, поскольку возможно наличие аргиназы и при: отсутствии всего цикла.
МОЧЕВИНА У ПОЗВОНОЧНЫХ
Позвоночные животные, выделяющие главным образом мочевину и обладающие для ее синтеза ферментами орнитинового цикла, представлены на рис. 10.14. Некоторое количество мочевины выделяют костистые рыбы, а у пластиножаберных, амфибий и млекопитающих это главный азотистый экскрет. У пластиножаберных (акул и скатов), а также у крабоядной лягушки и целаканта Latimeria мочевина задерживается в организме, играет
важную роль в саморегуляции и поэтому является ценным продуктом обмена. У пластиножаберных мочевина фильтруется в почечном клубочке, но ввиду ее значения для осморегуляции она не должна теряться с мочой; поэтому она возвращается в результате активной реабсорбции в канальцах. У амфибий дело обстоит иначе.

Мочевина фильтруется, и, кроме того, значительное количество ее добавляется к моче путем активной секреции в канальцах. Таким образом, и у пластиножаберных, и у амфибий имеет место активный канальцевый транспорт мочевины, но он идет у этих групп в разных направлениях. Очевидно, насосные механизмы здесь метаболически не идентичны, поскольку опыты с рядом близких друг к другу производных мочевины дают в обеих группаx животных различные результаты (табл. 10.5). Это превосходный пример того, как одна и та же физиологическая функция возникает в двух группах независимо, причем для достижения одной и той же цели (в данном случае для активного транспорта мочевины) не обязательно используются одинаковые механизмы.
У крабоядной лягушки, которая тоже сохраняет мочевину для осморегуляции, активной реабсорбции этого вещества в канальцах
не обнаружено (Schmidt-Nielsen, Lee, 1962). Моча образуется у нее медленно, и почечные канальцы весьма проницаемы для мочевины. Поэтому мочевина диффундирует из канальцевой жидкости

Рис. 10.14. Выделение азота на разных этапах филогенеза позвоночных. Линиям" окружены группы животных, которые выделяют соответственно аммиак, мочевину и мочевую кислоту в качестве основного экскрета. (В. Schmidt-Nielsen, 3972.)
обратно в кровь и оказывается в моче приблизительно в той же концентрации, что и в крови. Таким образом, лишь небольшие количества ее теряются с мочой.
Если у обычных лягушек происходит активная канальцевая секреция мочевины, то почему крабоядная лягушка не использует
Таблица 10.5
Мочевина активно транспортируется почечным канальцем акулы (активная реабсорбция) и лягушки (активная секреция). Но с тремя другими близкими веществами результаты получаются у этих двух видов животных совершенно различными. Это говорит о том, что клеточный механизм транспорта в их почках различен. (В. Schmidt-Nielsen, Rabinovitz, 1964)

такой насос, попросту поменяв его направление на обратное? На этот вопрос ответить нелегко, но, по-видимому, направление активного транспорта - консервативная физиологическая функция, изменить которую непросто. Как мы уже видели, и в коже лягушки, и в почке млекопитающего сохраняется направление активного транспорта хлористого натрия извне внутрь организма. Но в почке млекопитающего направленный внутрь обратный транспорт NaCl из канальцевой жидкости в организм используется в умножающей противоточной системе таким образом, что конечным результатом тем не менее оказывается концентрированная моча.
Обычное представление о выделении мочевины почкой млекопитающего состоит в том, что мочевина фильтруется в клубочке, а затем пассивно проходит по канальцам, хотя некоторая ее доля благодаря высокой способности к диффузии пассивно диффундирует обратно в кровь. Имеются, однако; убедительные доказательства того, что мочевина служит важным элементом умножающей противоточной системы и что способ выделения мочевины является существенным элементом функции почки у млекопитающих.
МОЧЕВИНА И МЕТАМОРФОЗ У АМФИБИЙ
Головастики лягушек и жаб выделяют главным образом аммиак; взрослые животные выделяют мочевину. У лягушки (Rana temporaria), жабы (Bufo bufo), тритона (Triturus uulgaris) и других амфибий при метаморфозе происходит четкий переход от выделения аммиака к экскреции мочевины. Однако южноафриканская шпорцевая лягушка (Xenopus), которая и во взрослом состоянии остается в воде, продолжает выделять аммиак и на этой: :тадии (табл. 10.6).
Переход к выделению мочевины во время метаморфоза у полуназемных амфибий связан; с заметным повышением активности всех ферментов орнитинового цикла в печени (Brown et al., 1959).
Таблица 10.6
Выделение аммиака у наземной жабы Bufo bufo и у полностью водной бесхвостой амфибии Xenopus laevis. Цифры указывают выделение свободного аммиака в процентах от общего количества выделяемых аммиака и мочевины на разных стадиях развития. (Munro, 1953 )

Интересно, что особи водной амфибии Xenopus, извлеченные:на несколько недель из воды, накапливают в крови и тканях мочевину. Накопление мочевины можно вызвать, поместив животных в 0,9%-ный раствор NaCl. Когда взрослых особей содержали вне воды, но в сыром мху (чтобы избежать обезвоживания), концентрация мочевины в крови увеличивалась в 10-.20 раз и достигала почти 100 ммоль/л. После возвращения животных в воду избыточная мочевина выделялась (Balinsky et al, 1961).
У группы особей Xenopus, которые в естественных условиях переживали летнюю засуху в иле около высохшего пруда, концентрация мочевины также была повышена в 15-20 раз. Среди ферментов, участвующих в синтезе мочевины, количество карбамоилфосфатсинтетазы, ответственной за первый этап синтеза (см. рис. 10.13), возросло приблизительно в шесть раз, но активность остальных ферментов цикла не изменилась. Возможно, что синтез карбамоилфосфата является этапом, лимитирующим скорость синтеза мочевины, и увеличение количества этого фермента, вероятно, удерживает аммиак плазмы на низком уровне, когда животные находятся вне воды (Balinsky et al., 1967).
МОЧЕВИНА У ДВОЯКОДЫШАЩИХ РЫБ
У африканской двоякодышащей рыбы Protopterus происходят совершенно такие же изменения, как у амфибий. В обычных условиях, когда такая рыба живет в воде, она выделяет много аммиака
(и некоторое количество мочевины), но когда в период засухи она находится в коконе в засохшем иле, то все азотистые отходы превращаются у нее в мочевину, которая накапливается в крови, где ее концентрация к концу трехлетнего пребывания рыбы в коконе может достигать 3% (500 ммоль/л) (Smith, 1959).
В печени африканской двоякодышащей рыбы обнаружены все пять ферментов орнитинового цикла (Janssens, Cohen, 1966). Уровни двух ферментов, лимитирующих скорость синтеза мочевины, сходны у этой рыбы и у головастика лягушки Rana catesbeiaпа и значительно ниже уровней, найденных у взрослых лягушек. Это согласуется с фактом преимущественного выделения аммиака двоякодышащей рыбой, когда она находится в воде. Однако было вычислено, что того количества ферментов орнитинового цикла, которое содержится в печени двоякодышащей рыбы, не находящейся в спячке, достаточно, чтобы обеспечить накопление мочевины, фактически наблюдаемое во время спячки (Forster, Goldstein, 1966).
У австралийской двоякодышащей рыбы Neoceratodus концентрации ферментов орнитинового цикла невелики, что согласуется с образом жизни этой рыбы: она пользуется легким только как: добавочным органом дыхания и может лишь недолго выживать на воздухе (о дыхании двоякодышащих рыб см. гл. 2). Синтез мочевины в срезах печени австралийской двоякодышащей рыбы идет в сто раз медленнее, чем у африканской. Это опять-таки согласуется с чисто водным образом жизни первой из них (Goldstein et al., 1967).
МОЧЕВАЯ КИСЛОТА
Выделение мочевой кислоты преобладает у насекомых, наземных улиток, большинства рептилий и у птиц. Все это - типично наземные животные, и образование у них мочевой кислоты можно рассматривать как эффективное приспособление, сберегающее воду при жизни на суше. Поскольку мочевая кислота и ее соли очень плохо растворимы в воде (растворимость ее составляет около 6 мг на 1 л воды), реабсорбция воды из мочи ведет к выпадению мочевой кислоты и ее солей в осадок.
МОЧЕВАЯ КИСЛОТА У ПТИЦ И НАСЕКОМЫХ
Полутвердая белая часть птичьего помета представляет собой мочу и состоит главным образом из мочевой кислоты; для выведения азотистых экскретов птицы расходуют очень мало воды, некоторых насекомых уменьшение потерь воды с мочой зашло так далеко, что они вообще не выделяют мочевую кислоту, а откладывают ее в разных частях организма, главным образом в
жировом теле. Поэтому для удаления конечных азотистых продуктов таким формам вода совсем не нужна (Kilby, 1963).
Было высказано предположение, что использование мочевой кислоты в качестве главного экскрета дает птицам еще одно преимущество. Поскольку для образования мочи им нужно мало воды, выделение мочевой кислоты, согласно этому предположению, уменьшает вес тела у летающих птиц. Но это соображение не убедительно, так как птицы, имеющие доступ к воде (как пресноводные, так и морские), часто выделяют большие количества жидкой мочи.
КЛЕЙДОИЧЕСКОЕ ЯЙЦО
Джозеф Нидхем предположил, что разница между теми позвоночными, которые вырабатывают мочевину (млекопитающие и амфибии), и теми, которые продуцируют мочевую кислоту (рептилии и птицы), связана прежде всего со способом размножения. Яйцо амфибии развивается в воде, а эмбрион млекопитающего - в жидкой среде в матке, где отходы метаболизма попадают в кровь матери. С другой стороны, эмбриональное развитие рептилий и птиц происходит в замкнутом, так называемом клейдоическом яйце, которое обменивается с внешней средой только газами, а все экскреты остаются внутри скорлупы. В клейдоическом яйце запас воды очень невелик, а аммиак, разумеется, слишком токсичен, чтобы эмбрион мог выносить его присутствие в больших количествах. Если бы вырабатывалась мочевина, она оставалась бы в яйце и накапливалась в растворенном состоянии. Между тем мочевая кислота может выпадать в осадок и тем самым по существу элиминироваться; это и происходит, когда она откладывается в виде кристаллов в аллантоисе, который, таким образом, служит эмбриональным мочевым пузырем.
МОЧЕВАЯ КИСЛОТА У РЕПТИЛИЙ
Ящерицы и змеи выделяют главным образом мочевую кислоту; многие черепахи выделяют смесь мочевой кислоты и мочевины, а крокодилы - главным образом аммиак (Cragg et al., 1961). Это соответствует общему представлению, что способ экскреции азота тесно связан с количеством доступной воды в окружающей среде.
Крокодилы и аллигаторы выделяют аммиак в моче, где главным катионом является NH4+, а главным анионом - НСО 3 - (Со-ulson et al., 1950; Goulson, Hernandez, 1955). Возможно, что присутствие в моче указанных ионов помогает этим пресноводным
животным лучше удерживать ионы Na + и С1 - , потеря которых с калом, кстати, тоже очень невелика.
Вряд ли можно сомневаться в тесной связи между средой обитания черепах и выделением ими азота. В табл. 10.7 приведен состав проб мочи от восьми видов черепах, полученных из Лондонского зоопарка. У видов с наиболее выраженным водным образом жизни выделяются значительные количества аммиака и мочевины и только следы мочевой кислоты; у наиболее сухопутных форм больше половины азота выводится в виде мочевой кислоты.
Таблица 10.7
Доля азота в моче различных черепах (в процентах от всего выделяемого азота). Формы, в наибольшей степени связанные с водой, почти не выделяют мочевой кислоты, но это вещество доминирует у наземных видов из засушливых областей. Moyle, 1949 )

Сведения о том, выделяют ли черепахи главным образом мочевину или же мочевую кислоту, противоречивы. Дело в том, что различаются не только виды, но и внутри одного вида одни особи могут выделять преимущественно мочевую кислоту, другие - преимущественно мочевину, третьи - смесь обоих веществ Khalil, Haggag, 1955). Даже одна и та же особь может с течением времени перейти от одного соединения к другому. Некоторое
количество выпавшей в осадок мочевой кислоты задерживается в клоаке, а жидкая часть мочи выводится наружу; это делает ненадежным определение образующейся мочевой кислоты путем анализа одной или нескольких проб мочи: при неполном опорожнении клоаки могут получиться совсем низкие цифры, а при такой ее эвакуации, когда выходит осадок, накопившийся за некоторое время, мочевой кислоты окажется слишком много.
У черепахи Testudo mauritanica переход от мочевины к мочевой кислоте и обратно находится, по-видимому, в прямой зависимости от температуры и содержания воды в организме. Выделение мочевой кислоты возрастает при неблагоприятном водном балансе, но механизм, управляющий этим сдвигом биохимической активности, неясен.
В главе 9 мы уже упоминали, что африканская лягушка Chiromantis xerampelina теряет воду через кожу очень медленно, примерно с той же скоростью, что и рептилии. Она сходна с рептилиями и тем, что выделяет в основном мочевую кислоту, а не мочевину, как это обычно свойственно взрослым амфибиям. Эта сенсационный факт, так как он противоречит общепринятому представлению о выделении азота у амфибий. Точность этого сообщения не вызывает сомнений, так как мочевую кислоту определяли в моче Chiromantis специфичным для этого вещества ферментативным методом, и было установлено, что она составляет до 60-75% сухого веса мочи (Loveridge, 1970).
Южноафриканская лягушка Phyllotnedusa sauvagii в этом отношении тоже сходна с рептилиями. Потеря воды через кожу составляет у нее величину того же порядка, что и у рептилий с их сухими покровами, а моча содержит много полутвердого осадка урата (Shoemaker et al., 1972). В форме урата у Phyllomedusa выделяется 80% общего азота, и повышенное потребление воды не изменяет интенсивности образования урата. Этот вид продолжает выделять главным образом мочевую кислоту даже при избытке воды. Когда лягушка нуждается в сохранении воды, экскреция мочевой кислоты (вместо мочевины) приобретает очень большое значение. Вычислено, что если бы экскреторным продуктом у этой лягушки была мочевина, то для образования мочи ей потребовалось бы около 60 мл воды в день на 1 кг веса тела. А между тем, благодаря тому что P. sauvagii выделяет мочевую кислоту, она теряет с мочой всего лишь 3,8 мл воды в день на 1 кг веса тела (Shoemaker, McClanahan, 1975).
АММИАК И ПОЧЕЧНАЯ ФУНКЦИЯ
Из всего сказанного выше может создаться впечатление, что аммиак выделяют главным образом водные животные, но это не совсем верно. Аммиак в норме содержится и в моче наземных животных, где он служит для регуляции рН мочи. Если моча становится кислой из-за выделения кислых продуктов обмена, для нейтрализации добавляется аммиак.
Избыток кислоты обычно образуется при белковом обмене, так как конечным продуктом окисления серусодержащей аминокислоты цистеина является серная кислота. Чем кислее моча, тем больше добавляется аммиака. Аммиак, используемый для нейтрализации кислой мочи, образуется в почках из аминокислоты глутамина. Почки содержат глутаминазу, и она имеется здесь специально для выработки аммиака. Поэтому аммиак в моче млекопитающего прямо не связан с тем аммиаком, который образуется в печени при дезаминировании аминокислот, и в этом смысле его не следует рассматривать как нормальный конечный продукт белкового обмена.
НУКЛЕИНОВЫЕ КИСЛОТЫ И ВЫДЕЛЕНИЕ АЗОТА
Нуклеиновые кислоты содержат две группы азотистых соединений: пурины (аденин и гуанин) и пиримидины (цитозин и тимин). У некоторых животных пурины выделяются в виде мочевой кислоты (которая и сама является пурином); у других животных пуриновая структура расщепляется до ряда промежуточных соединений или до аммиака, причем любое из этих веществ может выводиться из организма.
Метаболическое расщепление пуринов и выделение его конечных продуктов изучены не так тщательно, как обмен белкового 13ота. Важнейшие данные приведены в табл. 10.8. У птиц, наземных рептилий и насекомых пурины расщепляются до мочевой кислоты и последняя выводится из организма. Это те животные, у которых из аминного азота синтезируется мочевая кислота; очевидно, что для животного было бы бессмысленно синтезировать мочевую кислоту и в то же время обладать механизмами ее разложения. Поэтому нельзя ожидать дальнейшего распада пуринов у тех животных, у которых мочевая кислота - конечный продукт белкового обмена.
Среди млекопитающих человек, высшие обезьяны и далматский дог составляют особую группу: они выделяют мочевую кислоту, тогда как остальные млекопитающие выделяют аллантоин. Аллантоин образуется из мочевой кислоты путем одного превращения в присутствии фермента уриказы. У человека и высших обезьян нет этого фермента. Из-за своей малой растворимости мочевая кислота иногда откладывается в организме человека, вызывая припухлость суставов и очень мучительное заболевание -
Таблица 10.8
Азотистые конечные продукты пуринового обмена у разных животных. (Keilin, 1959)

подагру. Если бы у человека сохранился фермент уриказа, подагры не существовало бы.
Хотя далматский дог выделяет гораздо больше мочевой кислоты, чем другие собаки, это не следствие какого-то дефекта метаболизма. Печень всякой собаки содержит уриказу и вырабатывает некоторое количество аллантоина. Но у далматского дога имеется почечный дефект, препятствующий канальцевой реабсорбции мочевой кислоты (которая происходит у других млекопитающих, в том числе у человека); поэтому у дога мочевая кислота теряется с мочой быстрее, чем перерабатывается печенью в аллантоин (Yu et al., I960). Немало данных говорит о том, что мочевая кислота у далматского дога не только фильтруется в клубочке, но и экскретируется путем активного транспорта в канальцах (Keilin, 1959).
Пурины аденин и гуанин сходны по своей структуре с мочевой кислотой: они содержат одно шестичленное кольцо и одно пятичленное. Но пиримидины (цитозин и тимин) представляют собой одиночные шестичленные кольца, содержащие два атома азота. У высших позвоночных пиримидины расщепляются путем разрыва этого кольца с образованием одной молекулы аммиака и одной молекулы β-аминокислоты. Последняя метаболизируется затем по обычной схеме дезаминирования.
Самая поразительная черта обмена нуклеиновых кислот состоит в том, что "высшие" животные, перечисленные в начале табл. 10.8, полностью лишены ферментов, нужных для расщепления
пуринов. Среди "низших" животных мы находим все большее усложнение биохимических и ферментных систем, осуществляющих дальнейшее расщепление пуринов, так что самые "низшие" формы обладают наиболее полным ферментным аппаратом.
ДРУГИЕ АЗОТИСТЫЕ СОЕДИНЕНИЯ
У пауков главным экскретом является гуанин. По-видимому, он синтезируется из аминного азота, хотя весь путь его образования неизвестен. Некоторые пауки, в том числе птицеядные тарантулы, после приема пищи выделяют более 90% всего азота в форме гуанина (Peschen, 1939). У обыкновенного садового паука Epeira diadema идентификация гуанина была подтверждена весьма специфическим ферментативным методом (Vajropala, 1935).
Гуанин довольно часто встречается и у разнообразных других животных. Например, серебристый блеск рыбьей чешуи обусловлен отложением кристаллов гуанина. Садовая улитка Helix выделяет гуанин, но лишь в пределах около 20% общего количества экскретируемых пуринов, а остальные 80% составляет мочевая кислота. Возможно, что эта фракция - продукт обмена нуклеиновых кислот, а мочевая кислота образуется в результате белкового обмена.
Аминокислоты не занимают важного места среди продуктов азотистого обмена, но в небольших количествах они содержатся в моче многих животных. Казалось бы, животному выгоднее дезаминировать аминокислоту, выделять аммиак обычным путем и использовать образующуюся органическую кислоту в энергетическом обмене. Но поскольку аминокислоты играют лишь незначительную роль в выделении азота, этот вопрос не будет здесь обсуждаться.
ТЕОРИЯ РЕКАПИТУЛЯЦИИ
Обычно считали, что выделение азота у развивающегося куриного эмбриона изменяется во времени и проходит через ряд пиков: вначале основным продуктом является аммиак, затем мочевина и, наконец, мочевая кислота. Предполагалось, что такое развитие рекапитулирует этапы эволюции, которая у птиц заканчивается выделением мочевой кислоты. Как сообщалось, образование аммиака у куриного зародыша достигает максимума через 4 дня, мочевины - через 9 дней и мочевой кислоты - через 11 дней после начала инкубации (Baldwin, 1949).
Более новые работы говорят о том, что выделение азота у куриного эмбриона резко отличается от этой ранее описанной картины (Clark, Fischer, 1957). Все три главных экскреторных продукта- аммиак, мочевина и мочевая кислота - образуются и присутствуют с самого начала эмбрионального развития. К концу
периода инкубации мочевой кислоты становится намного больше, чем остальных двух продуктов. Однако количество мочевины и аммиака продолжает расти на протяжении всей инкубации, и ко времени вылупления оба вещества содержатся примерно в одинаковых количествах. К концу инкубации количество выделяемого азота достигает 40 мг, из которых 23% делятся поровну между мочевиной и аммиаком, а остальное представлено мочевой кислотой (рис. 10.15).

В чем причина расхождения в полученных результатах? Прежние данные могли быть менее точными из-за более примитивных методов анализа, но этим вряд ли можно объяснить наблюдавшиеся отдельные пики. Главная причина состоит просто в том, что результаты выражались в количествах каждого экскреторного продукта на единицу веса зародыша. А поскольку зародыш непрерывно и чем дальше, тем быстрее увеличивается в размерах,
то при делении количества каждого вещества на вес эмбриона создается искусственный пик.
На самом деле все три экскреторных продукта присутствуют с самого начала и на протяжении эмбрионального развития их становится постепенно все больше, но после 10-го дня инкубации количество аммиака возрастает незначительно. Мочевина, вырабатываемая зародышем, синтезируется не из азота аминокислот в цикле орнитина, а в результате воздействия аргиназы на аргинин (Eakin, Fisher, 1958). Таким образом, ни образование аммиака, ни синтез мочевины в курином эмбрионе не подтверждают представление о том, что онтогенез биохимических механизмов повторяет эволюционную историю выделения азота.
Мы рассмотрели разнообразные органы выделения и описали их общие особенности. Эти органы удаляют отходы метаболизма, помогают поддерживать нужные концентрации солей и других растворенных веществ и регулируют содержание воды в организме, тщательно сохраняя воду, если ее в организме мало, и выводя ее избыточные количества.
Убедительные данные показывают, однако, что это не всегда верно, Некоторые исследователи (например, Costa et al., 1968, 1974) сообщают об образовании газообразного азота у млекопитающих, получающих большие количества белка. Эти сведения должны изменить некоторые из наших представлений о белковом обмене и конечных азотистых продуктах.
По-гречески kleisto - замкнутый, от kleis - ключ.
Два описанных здесь вида лягушек обитают в сухих, полупустынных местностях. - Прим. ред.
Вопрос полный
Азотистый обмен
Азотистый обмен - совокупность химических превращений, реакций синтеза и распада азотистых соединений в организме; составная часть обмена веществ и энергии. Понятие «азотистый обмен» включает в себя белковый обмен (совокупность химических превращений в организме белков и продуктов их метаболизма), а также обмен пептидов, аминокислот , нуклеиновых кислот , нуклеотидов, азотистых оснований, аминосахаров (см. Углеводы), азотсодержащих липидов , витаминов , гормонов и других соединений, содержащих азот.
Организм животных и человека усвояемый азот получает с пищей, в которой основным источником азотистых соединений являются белки животного и растительного происхождения. Главным фактором поддержания азотистого равновесия - состояния А. о., при котором количество вводимого и выводимого азота одинаково, - служит адекватное поступление белка с пищей. В СССР суточная норма белка в питании взрослого человека принята равной 100 г , или 16 г азота белка, при расходе энергии 2500 ккал . Азотистый баланс (разность между количеством азота, который попадает в организм с пищей, и количеством азота, выводимого из организма с мочой, калом, потом) является показателем интенсивности А. о. в организме. Голодание или недостаточное по азоту питание приводят к отрицательному азотистому балансу, или азотистому дефициту, при котором количество азота, выводимого из организма, превышает количество азота, поступающего в организм с пищей. Положительный азотистый баланс, при котором вводимое с пищей количество азота превышает количество азота, выводимое из организма, наблюдается в период роста организма, при процессах регенерации тканей и т.д. Состояние А. о. в значительной степени зависит от качества пищевого белка, которое, в свою очередь, определяется его аминокислотным составом и прежде всего наличием незаменимых аминокислот.
Принято считать, что у человека и позвоночных животных А. о. начинается с переваривания азотистых соединений пищи в желудочно-кишечном тракте. В желудке происходит расщепление белков при участии пищеварительных протеолитических ферментов трипсина и гастриксина (см. Протеолиз ) с образованием полипептидов, олигопептидов и отдельных аминокислот. Из желудка пищевая масса поступает в двенадцатиперстную кишку и нижележащие отделы тонкой кишки, где пептиды подвергаются дальнейшему расщеплению, катализируемому ферментами сока поджелудочной железы трипсином, химотрипсином и карбоксипептидазой и ферментами кишечного сока аминопептидазами и дипептидазами (см. Ферменты ). Наряду с пептидами. в тонкой кишке расщепляются сложные белки (например, нуклеопротеины) и нуклеиновые кислоты. Существенный вклад в расщепление азотсодержащих биополимеров вносит и микрофлора кишечника. Олигопептиды, аминокислоты, нуклеотиды, нуклеозиды и др. всасываются в тонкой кишке, поступают в кровь и с ней разносятся по всему организму. Белки тканей организма в процессе постоянного обновления также подвергаются протеолизу под действием тканевых протсаз (пептидаз и катепсинов), а продукты распада тканевых белков попадают в кровь. Аминокислоты могут быть использованы для нового синтеза белков и других соединений (пуриновых и пиримидиновых оснований, нуклеотидов, порфиринов и т.д.), для получения энергии (например, посредством включения в цикл трикарбоновых кислот) или могут быть подвергнуты дальнейшей деградации с образованием конечных продуктов А. о., подлежащих выведению из организма.
Аминокислоты, поступающие в составе белков пищи, используются для синтеза белков органов и тканей организма. Они участвуют также в образовании многих других важных биологических соединений: пуриновых нуклеотидов (глутамин, глицин, аспарагиновая кислота) и пиримидиновых нуклеотидов (глутамин, аспарагиновая кислота), серотонина (триптофан), меланина (фенилалпнин, тирозин), гистамина (гистидин), адреналина, норадреналина, тирамина (тирозин), полиаминов (аргинин, метионин), холина (метионин), порфиринов (глицин), креатина (глицин, аргинин, метионин), коферментов, сахаров и полисахаридов, липидов и т.д. Важнейшей для организма химической реакцией, в которой участвуют практически все аминокислоты, является трансаминирование, заключающееся в обратимом ферментативном переносе a-аминогруппы аминокислот на a-углеродный атом кетокислот или альдегидов. Трансаминирование является принципиальной реакцией биосинтеза заменимых аминокислот в организме. Активность ферментов, катализирующих реакции трансаминирования, - аминотрансфераз - имеет большое клинико-диагностическое значение.
Деградация аминокислот может протекать по нескольким различным путям. Большинство аминокислот способно подвергаться декарбоксилированию при участии ферментов декарбоксилаз с образованием первичных аминов, которые затем могут окисляться в реакциях, катализируемых моноаминоксидазой или диаминоксидазой. При окислении биогенных аминов (гистамина, серотонина, тирамина, g-аминомасляной кислоты) оксидазами образуются альдегиды, подвергающиеся дальнейшим превращениям, и аммиак , основным путем дальнейшего метаболизма которого является образование мочевины.
Другим принципиальным путем деградации аминокислот является окислительное дезаминирование с образованием аммиака и кетокислот. Прямое дезаминирование L-аминокислот в организме животных и человека протекает крайне медленно, за исключением глутаминовой кислоты, которая интенсивно дезаминируется при участии специфического фермента глутаматдегидрогеназы. Предварительное трансаминирование почти всех a-аминокислот и дальнейшее дезаминирование образовавшейся глутаминовой кислоты на a-кетоглутаровую кислоту и аммиак является основным механизмом дезаминирования природных аминокислот.
Продуктом разных путей деградации аминокислот является аммиак, который может образовываться и в результате метаболизма других азотсодержащих соединений (например, при дезаминировании аденина, входящего в состав никотинамидадениндинуклеотида - НАД). Основным путем связывания и нейтрализации токсичного аммиака у уреотелических животных (животные, у которых конечным продуктом А. о, является мочевина) служит так называемый цикл мочевины (синоним: орнитиновый цикл, цикл Кребса - Гензелейта), протекающий в печени. Он представляет собой циклическую последовательность ферментативных реакций, в результате которой из молекулы аммиака или амидного азота глутамина, аминогруппы аспарагановой кислоты и диоксида углерода осуществляется синтез мочевины. При ежедневном потреблении 100 г белка суточное выведение мочевины из организма составляет около 30 г . У человека и высших животных существует еще один путь нейтрализации аммиака - синтез амидов дикарбоновых кислот аспарагана и глутамина из соответствующих аминокислот. У урикотелических животных (рептилии, птицы) конечным продуктом А. о. является мочевая кислота.
В результате расщепления нуклеиновых кислот и нуклеопротеинов в желудочно-кишечном тракте образуются нуклеотиды и нуклеозиды. Олиго- и моно-нуклеотиды при участии различных ферментов (эстераз, нуклеотидаз, нуклеозидаз, фосфорилаз) превращаются затем в свободные пуриновые и пиримидиновые основания.
Дальнейший путь деградации пуриновых оснований аденина и гуанина состоит в их гидролитическом дезаминировании под влиянием ферментов аденазы и гуаназы с образованием соответственно гипоксантина (6-оксипурина) и ксантина (2,6-диоксипурина), которые затем превращаются в мочевую кислоту в реакциях, катализируемых ксантиноксидазой. Мочевая кислота - один из конечных продуктов А. о. и конечный продукт обмена пуринов у человека - выводится из организма с мочой. У большинства млекопитающих имеется фермент уриказа, который катализирует превращение мочевой кислоты в экскретируемый аллантоин.
Деградация пиримидиновых оснований (урацила, тимина) состоит в их восстановлении с образованием дигидропроизводных и последующем гидролизе, в результате которого из урацила образуется b-уреидопропионовая кислота, а из нее - аммиак, диоксид углерода и b-аланин, а из тимина - b-аминоизомасляная кислота, диоксид углерода и аммиак. Диоксид углерода и аммиак могут далее включаться в мочевину через цикл мочевины, а b-аланин участвует в синтезе важнейших биологически активных соединений - гистидинсодержащих дипептидов карнозина (b-аланил-L-гистидина) и анзерина (b-аланил-N-метил-L-гистидина), обнаруживаемых в составе экстрактивных веществ скелетных мышц, а также в синтезе пантотеновой кислоты и кофермента А.
Т.о., разнообразные превращения важнейших азотистых соединений организма связаны между собой в единый обмен. Сложный процесс А. о. регулируется на молекулярном, клеточном и тканевом уровнях. Регуляция А. о. в целом организме направлена на приспособление интенсивности А. о. к изменяющимся условиям окружающей и внутренней среды и осуществляется нервной системой как непосредственно, так и путем воздействия на железы внутренней секреции.
У здоровых взрослых людей содержание азотистых соединений в органах, тканях, биологических жидкостях находится на относительно постоянном уровне. Избыток азота, поступившего с пищей, выводится с мочой и калом, а при недостатке азота в пище нужды организма в нем могут покрываться за счет использования азотистых соединений тканей тела. При этом состав мочи изменяется в зависимости от особенностей А. о. и состояния азотистого баланса. В норме при неизменном режиме питания и относительно стабильных условиях окружающей среды из организма выделяется постоянное количество конечных продуктов А. о., а развитие патологических состояний приводит к его резкому изменению. Значительные изменения экскреции азотистых соединений с мочой, в первую очередь экскреции мочевины, могут наблюдаться и при отсутствии патологии в случае существенного изменения режима питания (например, при изменении количества потребляемого белка), причем концентрация остаточного азота (см. Азот остаточный ) в крови меняется незначительно.
При исследовании А. о. необходимо учитывать количественный и качественный состав принимаемой пищи, количественный и качественный состав азотистых соединений, выделяемых с мочой и калом и содержащихся в крови. Для исследования А. о. применяют азотистые вещества, меченные радионуклидами азота, фосфора, углерода, серы, водорода, кислорода, и наблюдают за миграцией метки и включением ее в состав конечных продуктов А. о. Широко используют меченые аминокислоты, например 15 N-глицин, которые вводят в организм с пищей или непосредственно в кровь. Значительная часть меченого азота глицина пищи выводится в составе мочевины с мочой, а другая часть метки попадает в тканевые белки и выводится из организма крайне медленно. Проведение исследования А. о. необходимо для диагностики многих патологических состояний и контроля за эффективностью лечения, а также при разработке рациональных схем питания, в т.ч. лечебного (см. Питание лечебное ).
Патологию А. о. (вплоть до очень значительной) вызывает белковая недостаточность. Ее причиной может стать общее недоедание, продолжительный дефицит белка или незаменимых аминокислот в рационе, недостаток углеводов и жиров, обеспечивающих энергией процессы биосинтеза белка в организме. Белковая недостаточность может быть обусловлена преобладанием процессов распада белков над их синтезом не только в результате алиментарного дефицита белка и других важнейших пищевых веществ, но и при тяжелой мышечной работе, травмах, воспалительных и дистрофических процессах, ишемии, инфекции, обширных ожогах, дефекте трофической функции нервной системы, недостаточности гормонов анаболического действия (гормона роста, половых гормонов, инсулина), избыточном синтезе или избыточном поступлении извне стероидных гормонов и т.п. Нарушение усвоения белка при патологии желудочно-кишечного тракта (ускоренная эвакуация пищи из желудка, гипо- и анацидные состояния, закупорка выводного протока поджелудочной железы, ослабление секреторной функции и усиление моторики тонкой кишки при энтеритах и энтероколитах, нарушение процесса всасывания в тонкой кишке и др.) также может приводить к белковой недостаточности. Белковая недостаточность ведет к дискоординации А. о. и характеризуется резко выраженным отрицательным азотистым балансом.
Известны случаи нарушения синтеза определенных белков (см. Иммунопатология , Ферментопатии ), а также генетически обусловленного синтеза аномальных белков, например при гемоглобинопатиях , миеломной болезни (см. Парапротеинемические гемобластозы ) и др.
Патология А. о., заключающаяся в нарушении обмена аминокислот, часто связана с аномалиями процесса трансаминирования: уменьшением активности аминотрансфераз при гипо- или авитаминозах В 6 , нарушением синтеза этих ферментов, недостатком кетокислот для трансаминирования в связи с угнетением цикла трикарбоновых кислот при гипоксии и сахарном диабете и т.д. Снижение интенсивности трансаминирования приводит к угнетению дезаминирования глутаминовой кислоты, а оно, в свою очередь, - к повышению доли азота аминокислот в составе остаточного азота крови (гипераминоацидемии), общей гиперазотемии и аминоацидурии. Гипераминоацидемия, аминоацидурия и общая азотемия характерны для многих видов патологии А. о. При обширных поражениях печени и других состояниях, связанных с массивным распадом белка в организме, нарушаются процессы дезаминирования аминокислот и образования мочевины таким образом, что возрастают концентрация остаточного азота и содержание в нем азота аминокислот на фоне снижения относительного содержания в остаточном азоте азота мочевины (так называемая продукционная азотемия).
Продукционная азотемия, как правило, сопровождается выведением избытка аминокислот с мочой, поскольку даже в случае нормального функционирования почек фильтрация аминокислот в почечных клубочках происходит интенсивнее, чем их реабсорбция в канальцах. Заболевания почек, обтурация мочевых путей, нарушение почечного кровообращения приводят к развитию ретенционной азотемии, сопровождающейся нарастанием концентрации остаточного азота в крови за счет повышения содержания в крови мочевины (см. Почечная недостаточность ). Обширные раны, тяжелые ожоги, инфекции, повреждения трубчатых костей, спинного и головного мозга, гипотиреоз, болезнь Иценко - Кушинга и многие другие тяжелые заболевания сопровождаются аминоацидурией. Она характерна и для патологических состояний, протекающих с нарушением процессов реабсорбции в почечных канальцах: болезни Вильсона - Коновалова (см. Гепатоцеребральная дистрофия ), нефронофтизе Фанкони (см. Рахитоподобные болезни ) и др. Эти болезни относятся к многочисленным генетически обусловленным нарушениям А. о. Избирательное нарушение реабсорбции цистина и цистинурия с генерализованным нарушением обмена цистина на фоне общей аминоацидурии сопровождает так называемый цистиноз. При этом заболевании кристаллы цистина откладываются в клетках ретикулоэндотелиальной системы. Наследственное заболевание фенилкетонурия характеризуется нарушением превращения фенилаланина в тирозин в результате генетически обусловленной недостаточности фермента фенилаланин - 4-гидроксилазы, что вызывает накопление в крови и моче непревращенного фенилаланина и продуктов его обмена - фенилпировиноградной и фенилуксусной кислот. Нарушение превращений этих соединений характерно и для вирусного гепатита.
Тирозинемию, тирозинурию и тирозиноз отмечают при лейкозах, диффузных заболеваниях соединительной ткани (коллагенозах) и других патологических состояниях. Они развиваются вследствие нарушения трансаминирования тирозина. Врожденная аномалия окислительных превращений тирозина лежит в основе алкаптонурии, при которой в моче накапливается непревращенный метаболит этой аминокислоты - гомогентизиновая кислота. Нарушения пигментного обмена при гипокортицизме (см. Надпочечники ) связаны с угнетением превращения тирозина в меланин вследствие ингибирования фермента тирозиназы (полное выпадение синтеза этого пигмента характерно для врожденной аномалии пигментации - альбинизма).
При хроническом гепатите, сахарном диабете, остром лейкозе, хроническом миело- и лимфолейкозе, лимфогранулематозе, ревматизме и склеродермии нарушается обмен триптофана и его метаболиты 3-оксикинуренин, ксантуреновая и 3-оксиантраниловая кислоты, обладающие токсическими свойствами, накапливаются в крови. К патологии А. о. относятся и состояния, связанные с нарушением выделения почками креатинина и накоплением его в крови. Усиление экскреции креатинина сопровождает гиперфункцию щитовидной железы, а снижение экскреции креатинина при повышенном выведении креатина - гипотиреоз.
При массивном распаде клеточных структур (голодание, тяжелая мышечная работа, инфекции и др.) отмечают патологическое нарастание концентрации остаточного азота за счет увеличения относительного содержания в ней азота мочевой кислоты (в норме концентрация мочевой кислоты в крови не превышает - 0,4 ммоль/л ).
В пожилом возрасте снижаются интенсивность и объем синтеза белка за счет непосредственного угнетения биосинтетической функции организма и ослабления его способности усваивать аминокислоты пищи; развивается отрицательный азотистый баланс. Нарушения обмена пуринов у людей пожилого возраста приводят к накоплению и отложению в мышцах, суставах и хрящах солей мочевой кислоты - уратов. Коррекция нарушений А. о. в пожилом возрасте может быть осуществлена за счет специальных диет, содержащих полноценные животные белки, витамины и микроэлементы, с ограниченным содержанием пуринов.
Азотистый обмен у детей отличается рядом особенностей, в частности положительным азотистым балансом как необходимым условием роста. Интенсивность процессов А. о. на протяжении роста ребенка подвергается изменениям, особенно ярко выраженным у новорожденных и детей раннего возраста. В течение первых 3-х дней жизни азотистый баланс отрицателен, что объясняется недостаточным поступлением белка с пищей. В этот период обнаруживается транзиторное повышение концентрации остаточного азота в крови (так называемая физиологическая азотемия), иногда достигающее 70 ммоль/л ; к концу 2-й нед.
жизни концентрация остаточного азота снижается до уровня, отмечаемого у взрослых. Количество выделяемого почками азота нарастает в течение первых 3-х дней жизни, после чего снижается и вновь начинает увеличиваться со 2-й нед. жизни параллельно возрастающему количеству пищи.
Наиболее высокая усвояемость азота в организме ребенка наблюдается у детей первых месяцев жизни. Азотистый баланс заметно приближается к равновесию в первые 3-6 мес. жизни, хотя и остается положительным. Интенсивность белкового обмена у детей достаточно высока - у детей 1-го года жизни обновляется около 0,9 г белка на 1 кг массы тела в сутки, в 1-3 года - 0,8 г/кг/ сут., у детей дошкольного и школьного возраста - 0,7 г/кг/ сут.
Средние величины потребности в незаменимых аминокислотах, по данным ФАО ВОЗ (1985), у детей в 6 раз больше, чем у взрослых (незаменимой аминокислотой для детей в возрасте до 3 мес. является цистин, а до 5 лет - и гистидин). Более активно, чем у взрослых, протекают у детей процессы трансаминирования аминокислот. Однако в первые дни жизни у новорожденных из-за относительно низкой активности некоторых ферментов отмечаются гипераминоацидемия и физиологическая аминоацидурия в результате функциональной незрелости почек. У недоношенных, кроме того, имеет место аминоацидурия перегрузочного типа, т.к. содержание свободных аминокислот в плазме их крови выше, чем у доношенных детей. На первой неделе жизни азот аминокислот составляет 3-4% общего азота мочи (по некоторым данным, до 10%), и лишь к концу 1-го года жизни его относительное содержание снижается до 1%. У детей 1-го года жизни выведение аминокислот в расчете на 1 кг массы тела достигает величин выведения их у взрослого человека, экскреция азота аминокислот, достигающая у новорожденных 10 мг/кг массы тела, на 2-м году жизни редко превышает 2 мг/кг массы тела. В моче новорожденных повышено (по сравнению с мочой взрослого человека) содержание таурина, треонина, серина, глицина, аланина, цистина, лейцина, тирозина, фенилаланина и лизина. В первые месяцы жизни в моче ребенка обнаруживаются также этаноламин и гомоцитруллин. В моче детей 1-го года жизни преобладают аминокислоты пролин и [гидр]оксипролин.
Исследования важнейших азотистых компонентов мочи у детей показали, что соотношение мочевой кислоты, мочевины и аммиака в процессе роста существенно изменяется. Так, первые 3 мес. жизни характеризуются наименьшим содержанием в моче мочевины (в 2-3 раза меньше, чем у взрослых) и наибольшей экскрецией мочевой кислоты. Дети в первые три месяца жизни выделяют 28,3 мг/кг массы тела мочевой кислоты, а взрослые - 8,7 мг/кг . Относительно высокая экскреция у детей первых месяцев жизни мочевой кислоты способствует иногда развитию мочекислого инфаркта почек. Количество мочевины в моче нарастает у детей в возрасте от 3 до 6 месяцев, а содержание мочевой кислоты в это время снижается. Содержание аммиака в моче детей в первые дни жизни невелико, но затем резко возрастает и держится на высоком уровне на протяжении всего 1-го года жизни.
Характерной особенностью А. о. у детей является физиологическая креатинурия. Креатин обнаруживается еще в амниотической жидкости; в моче он определяется в количествах, превышающих содержание креатина в моче взрослых, начиная с периода новорожденности и до периода полового созревания. Суточная экскреция креатинина (дегидроксилированного креатина) с возрастом увеличивается, в то же время по мере нарастания массы тела ребенка относительное содержание азота креатинина мочи снижается. Количество креатинина, выводимого с мочой за сутки, у доношенных новорожденных составляет 10-13 мг/кг , у недоношенных 3 мг/кг , у взрослых не превышает 30 мг/кг.
При выявлении в семье врожденного нарушения А. о. необходимо проведение
Вопрос короткр
Выделение конечных продуктов азотистого обмена
Мочевая кислота является одним из наиболее важных конечных продуктов азотистого обмена у человека. В норме ее концентрация в сыворотке крови у мужчин составляет 0,27- 0,48 ммоль-л1, у женщин 0,18-0,38 ммоль-л-1; суточная экскреция с мочой колеблется от 2,3 до 4,5 ммоль (400-750 мг). У человека экскретируется мочевая кислота, у многих млекопитающих имеется фермент уриказа, которая окисляет мочевую кислоту до аллантоина. В теле здорового человека в сутки образование и выделение мочевой кислоты составляет от 500 до 700 мг. Большая часть мочевой кислоты (до 80 %) образуется в результате метаболизма эндогенных нуклеиновых кислот, лишь около 20 % связано с пуринами, поступающими с пищей. Почки за сутки экскретируют около 500 мг мочевой кислоты, 200 мг удаляются через желудочно-кишечный тракт.
Функциональные протеинурии. К функциональным протеинуриям, точные процессы возникновения которых не определены, причисляются относящаяся к вертикальному положению тела, идиопатическая непостоянное, выделение белка с мочой натуги, горячечное появление белка в моче, а также выделение белка с мочой приожирении.
Для ортостатической протеинурии свойственно появление полипептида в urina при длительном бездействии или расхаживании с скорым его исчезновением при изменении позы тела на перпендикулярное. Появление белка в моче в большинстве случаев не превосходит один г/сутки, является клубочковой и невыборочной, процедура ее появления неотчётлива. Чаще она отмечается в юношеском возрасте, у половины больных вылечивается через некоторое время. Механизм формирования, пожалуй, объединен с ненормально обострённым ответом почечного кровообращения на изменение размещения туловища.
Определение ортостатической протеинурии ставят при комбинировании ниженазванных условий:
Возраст пациентов 13-двадцать лет;
Закрытый тип появления белка в моче, неимение иных признаков нарушения почек (перестроений уринозного осадка, увеличения давления, которое кровь, находящаяся в артерии, оказывает на ее стенку, видоизменений сосудов внутренней поверхности глазного яблока);
Всего лишь ортостатический ход появления белка в моче, когда в изучениях урины, забранных после нахождения обследуемого в положении лежа (в том числе наутро до вставания с постели), протеин отсутствует.
Для доказательства этого диагноза нужно выполнить относящуюся к вертикальному положению туловища проверку. Для этого урину собирают утром перед вставанием с кровати, затем после некоторого нахождения в перпендикулярном положении (движение с палкой за спиной для развёртывания позвоночного столба). Диагностика передаёт еще более точные итоги, когда утренняя (ночная) доля urina сливается (так как в vesica urinaria возможна остаточная урина), а начальная порция подбирается после недолгого присутствия пациента в лежачем положении.
В молодом возрасте возможна в свою очередь первичное непостоянное появление белка в моче, устанавливаемая у здоровых индивидуумов при врачебном исследовании и пропадающая при следующих анализах мочи.
Протеинурия напряжения выявляется у двадцати процентов лиц в здоровом состоянии (даже спортсменов) после сильной физической нагрузки. Белок выявляетсяв начальной подготовленной порции урины. выделение белка с мочой характером связанным с патологией канальцев. Предполагается, что алгоритм появления белка в моче объединен с рекомбинацией тока крови и относительной ишемией проксимальных участков нефрона.
Лихорадочное появление белка в моче встречается при сильных горячих состояниях, в частности у лиц детского возраста и лиц старческого возраста. Лихорадочное выделение белка с мочой имеет преимущественно гломерулярный ход. Процессы этого вида протеинурии мало изучены, изучается возможная важность увеличения клубочковой фильтрации наравне с недолговечным поражением клубочкового фильтра защитными комплексами.
Выделение белка с мочой при патологической избыточной массе тела. Выделение белка с мочой нередко наблюдается при анормальном отложении жира в теле. (вес тела более 115 килограммов). По данным J.P.Domfeld (1989), среди одной тысячи пациентов с патологическим отложением жира в теле. у 420 диагностировано выделение белка с мочой без перерождений отстоя урины; показаны и прецеденты нефротического синдрома. Предполагается, что в первопричине формирования такой протеинурии располагаются искажения кровообращения скопления сильно фенестрированных капилляров (повышение давления в группе капилляров почечного тельца, повышенная скорость фильтрации), связанные с повышением при патологической избыточной массе тела. концентрации полипептидного гормона, вырабатываемого почкой вследствие понижения кровяного давления и гипертензин, которая при голодании убавляется. При снижении веса, а также при лечении замедлителями АПФ выделение белка с мочой может убавляться, а также утрачиваться.
Вдобавок, протеинурия может иметь внепочечное начало. При присутствии лейкоцитов в анализе мочи и особенно появление крови в моче удостоверяющая реакция на полипептид может быть итогом распада форменных элементов крови при длительном стоянии урины, в данной обстановке ненормальным представляется появление белка в моче превышающее, 0,3 грамма/сут. Осадочные полипептидные проверки способны давать ложноположительные результаты при присутствии в urina йодосодержащих контрастных материалов, немалого числа сходных препаратов Penicillinum а также лекарственного вещества из группы полусинтепнческих беталактамовых антибиотиков, продуктов метаболизма сульфаниламидных препаратов.
Похожая информация.
Азотистый обмен - совокупность химических превращений, реакций синтеза и распада азотистых соединений в организме; составная часть обмена веществ и энергии. Понятие «азотистый обмен» включает в себя белковый обмен (совокупность химических превращений в организме белков и продуктов их метаболизма), а также обмен пептидов, аминокислот , нуклеиновых кислот , нуклеотидов, азотистых оснований, аминосахаров (см. Углеводы), азотсодержащих липидов , витаминов , гормонов и других соединений, содержащих азот.
Организм животных и человека усвояемый азот получает с пищей, в которой основным источником азотистых соединений являются белки животного и растительного происхождения. Главным фактором поддержания азотистого равновесия - состояния А. о., при котором количество вводимого и выводимого азота одинаково, - служит адекватное поступление белка с пищей. В СССР суточная норма белка в питании взрослого человека принята равной 100 г , или 16 г азота белка, при расходе энергии 2500 ккал . Азотистый баланс (разность между количеством азота, который попадает в организм с пищей, и количеством азота, выводимого из организма с мочой, калом, потом) является показателем интенсивности А. о. в организме. Голодание или недостаточное по азоту питание приводят к отрицательному азотистому балансу, или азотистому дефициту, при котором количество азота, выводимого из организма, превышает количество азота, поступающего в организм с пищей. Положительный азотистый баланс, при котором вводимое с пищей количество азота превышает количество азота, выводимое из организма, наблюдается в период роста организма, при процессах регенерации тканей и т.д. Состояние А. о. в значительной степени зависит от качества пищевого белка, которое, в свою очередь, определяется его аминокислотным составом и прежде всего наличием незаменимых аминокислот.
Принято считать, что у человека и позвоночных животных А. о. начинается с переваривания азотистых соединений пищи в желудочно-кишечном тракте. В желудке происходит расщепление белков при участии пищеварительных протеолитических ферментов трипсина и гастриксина (см. Протеолиз ) с образованием ептидов, олигопептидов и отдельных аминокислот. Из желудка пищевая масса поступает в двенадцатиперстную кишку и нижележащие отделы тонкой кишки, где пептиды подвергаются дальнейшему расщеплению, катализируемому ферментами сока поджелудочной железы трипсином, химотрипсином и карбоксипептидазой и ферментами кишечного сока аминопептидазами и дипептидазами (см. Ферменты ). Наряду с пептидами. в тонкой кишке расщепляются сложные белки (например, нуклеопротеины) и нуклеиновые кислоты. Существенный вклад в расщепление азотсодержащих биополимеров вносит и микрофлора кишечника. Олигопептиды, аминокислоты, нуклеотиды, нуклеозиды и др. всасываются в тонкой кишке, поступают в кровь и с ней разносятся по всему организму. Белки тканей организма в процессе постоянного обновления также подвергаются протеолизу под действием тканевых протсаз (пептидаз и катепсинов), а продукты распада тканевых белков попадают в кровь. Аминокислоты могут быть использованы для нового синтеза белков и других соединений (пуриновых и пиримидиновых оснований, нуклеотидов, порфиринов и т.д.), для получения энергии (например, посредством включения в цикл трикарбоновых кислот) или могут быть подвергнуты дальнейшей деградации с образованием конечных продуктов А. о., подлежащих выведению из организма.
Аминокислоты, поступающие в составе белков пищи, используются для синтеза белков органов и тканей организма. Они участвуют также в образовании многих других важных биологических соединений: пуриновых нуклеотидов (глутамин, глицин, аспарагиновая кислота) и пиримидиновых нуклеотидов (глутамин, аспарагиновая кислота), серотонина (триптофан), меланина (фенилалпнин, тирозин), гистамина (гистидин), адреналина, норадреналина, тирамина (тирозин), полиаминов (аргинин, метионин), холина (метионин), порфиринов (глицин), креатина (глицин, аргинин, метионин), коферментов, сахаров и полисахаридов, липидов и т.д. Важнейшей для организма химической реакцией, в которой участвуют практически все аминокислоты, является трансаминирование, заключающееся в обратимом ферментативном переносе a -аминогруппы аминокислот на a -углеродный атом кетокислот или альдегидов. Трансаминирование является принципиальной реакцией биосинтеза заменимых аминокислот в организме. Активность ферментов, катализирующих реакции трансаминирования, - аминотрансфераз - имеет большое клинико-диагностическое значение.
Деградация аминокислот может протекать по нескольким различным путям. Большинство аминокислот способно подвергаться декарбоксилированию при участии ферментов декарбоксилаз с образованием первичных аминов, которые затем могут окисляться в реакциях, катализируемых моноаминоксидазой или диаминоксидазой. При окислении биогенных аминов (гистамина, серотонина, тирамина, g -аминомасляной кислоты) оксидазами образуются альдегиды, подвергающиеся дальнейшим превращениям, и аммиак , основным путем дальнейшего метаболизма которого является образование мочевины.
Другим принципиальным путем деградации аминокислот является окислительное дезаминирование с образованием аммиака и кетокислот. Прямое дезаминирование L-аминокислот в организме животных и человека протекает крайне медленно, за исключением глутаминовой кислоты, которая интенсивно дезаминируется при участии специфического фермента глутаматдегидрогеназы. Предварительное трансаминирование почти всех a -аминокислот и дальнейшее дезаминирование образовавшейся глутаминовой кислоты на a -кетоглутаровую кислоту и аммиак является основным механизмом дезаминирования природных аминокислот.
Продуктом разных путей деградации аминокислот является аммиак, который может образовываться и в результате метаболизма других азотсодержащих соединений (например, при дезаминировании аденина, входящего в состав никотинамидадениндинуклеотида - НАД). Основным путем связывания и нейтрализации токсичного аммиака у уреотелических животных (животные, у которых конечным продуктом А. о, является мочевина) служит так называемый цикл мочевины (синоним: орнитиновый цикл, цикл Кребса - Гензелейта), протекающий в печени. Он представляет собой циклическую последовательность ферментативных реакций, в результате которой из молекулы аммиака или амидного азота глутамина, аминогруппы аспарагановой кислоты и диоксида углерода осуществляется синтез мочевины. При ежедневном потреблении 100 г белка суточное выведение мочевины из организма составляет около 30 г . У человека и высших животных существует еще один путь нейтрализации аммиака - синтез амидов дикарбоновых кислот аспарагана и глутамина из соответствующих аминокислот. У урикотелических животных (рептилии, птицы) конечным продуктом А. о. является мочевая кислота.
В результате расщепления нуклеиновых кислот и нуклеопротеинов в желудочно-кишечном тракте образуются нуклеотиды и нуклеозиды. Олиго- и моно-нуклеотиды при участии различных ферментов (эстераз, нуклеотидаз, нуклеозидаз, фосфорилаз) превращаются затем в свободные пуриновые и пиримидиновые основания.
Дальнейший путь деградации пуриновых оснований аденина и гуанина состоит в их гидролитическом дезаминировании под влиянием ферментов аденазы и гуаназы с образованием соответственно гипоксантина (6-оксипурина) и ксантина (2,6-диоксипурина), которые затем превращаются в мочевую кислоту в реакциях, катализируемых ксантиноксидазой. Мочевая кислота - один из конечных продуктов А. о. и конечный продукт обмена пуринов у человека - выводится из организма с мочой. У большинства млекопитающих имеется фермент уриказа, который катализирует превращение мочевой кислоты в экскретируемый аллантоин.
Деградация пиримидиновых оснований (урацила, тимина) состоит в их восстановлении с образованием дигидропроизводных и последующем гидролизе, в результате которого из урацила образуется b -уреидопропионовая кислота, а из нее - аммиак, диоксид углерода и b -аланин, а из тимина - b -аминоизомасляная кислота, диоксид углерода и аммиак. Диоксид углерода и аммиак могут далее включаться в мочевину через цикл мочевины, а b -аланин участвует в синтезе важнейших биологически активных соединений - гистидинсодержащих дипептидов карнозина (b -аланил-L-гистидина) и анзерина (b -аланил-N-метил-L-гистидина), обнаруживаемых в составе экстрактивных веществ скелетных мышц, а также в синтезе пантотеновой кислоты и кофермента А.
Т.о., разнообразные превращения важнейших азотистых соединений организма связаны между собой в единый обмен. Сложный процесс А. о. регулируется на молекулярном, клеточном и тканевом уровнях. Регуляция А. о. в целом организме направлена на приспособление интенсивности А. о. к изменяющимся условиям окружающей и внутренней среды и осуществляется нервной системой как непосредственно, так и путем воздействия на железы внутренней секреции.
У здоровых взрослых людей содержание азотистых соединений в органах, тканях, биологических жидкостях находится на относительно постоянном уровне. Избыток азота, поступившего с пищей, выводится с мочой и калом, а при недостатке азота в пище нужды организма в нем могут покрываться за счет использования азотистых соединений тканей тела. При этом состав мочи изменяется в зависимости от особенностей А. о. и состояния азотистого баланса. В норме при неизменном режиме питания и относительно стабильных условиях окружающей среды из организма выделяется постоянное количество конечных продуктов А. о., а развитие патологических состояний приводит к его резкому изменению. Значительные изменения экскреции азотистых соединений с мочой, в первую очередь экскреции мочевины, могут наблюдаться и при отсутствии патологии в случае существенного изменения режима питания (например, при изменении количества потребляемого белка), причем концентрация остаточного азота (см. Азот остаточный ) в крови меняется незначительно.
При исследовании А. о. необходимо учитывать количественный и качественный состав принимаемой пищи, количественный и качественный состав азотистых соединений, выделяемых с мочой и калом и содержащихся в крови. Для исследования А. о. применяют азотистые вещества, меченные радионуклидами азота, фосфора, углерода, серы, водорода, кислорода, и наблюдают за миграцией метки и включением ее в состав конечных продуктов А. о. Широко используют меченые аминокислоты, например 15 N-глицин, которые вводят в организм с пищей или непосредственно в кровь. Значительная часть меченого азота глицина пищи выводится в составе мочевины с мочой, а другая часть метки попадает в тканевые белки и выводится из организма крайне медленно. Проведение исследования А. о. необходимо для диагностики многих патологических состояний и контроля за эффективностью лечения, а также при разработке рациональных схем питания, в т.ч. лечебного (см. Питание лечебное ).
Патологию А. о. (вплоть до очень значительной) вызывает белковая . Ее причиной может стать общее недоедание, продолжительный дефицит белка или незаменимых аминокислот в рационе, недостаток углеводов и жиров, обеспечивающих энергией процессы биосинтеза белка в организме. Белковая может быть обусловлена преобладанием процессов распада белков над их синтезом не только в результате алиментарного дефицита белка и других важнейших пищевых веществ, но и при тяжелой мышечной работе, травмах, воспалительных и дистрофических процессах, ишемии, инфекции, обширных ах, дефекте трофической функции нервной системы, недостаточности гормонов анаболического действия (гормона роста, половых гормонов, инсулина), избыточном синтезе или избыточном поступлении извне стероидных гормонов и т.п. Нарушение усвоения белка при патологии желудочно-кишечного тракта (ускоренная эвакуация пищи из желудка, гипо- и анацидные состояния, закупорка выводного протока поджелудочной железы, ослабление секреторной функции и усиление моторики тонкой кишки при энтеритах и энтероколитах, нарушение процесса всасывания в тонкой кишке и др.) также может приводить к белковой недостаточности. Белковая ведет к дискоординации А. о. и характеризуется резко выраженным отрицательным азотистым балансом.
Известны случаи нарушения синтеза определенных белков (см. Иммунопатология , Ферментопатии ), а также генетически обусловленного синтеза аномальных белков, например при гемоглобинопатиях , миеломной болезни (см. Парапротеинемические гемобластозы ) и др.
Патология А. о., заключающаяся в нарушении обмена аминокислот, часто связана с аномалиями процесса трансаминирования: уменьшением активности аминотрансфераз при гипо- или авитаминозах В 6 , нарушением синтеза этих ферментов, недостатком кетокислот для трансаминирования в связи с угнетением цикла трикарбоновых кислот при гипоксии и сахарном е и т.д. Снижение интенсивности трансаминирования приводит к угнетению дезаминирования глутаминовой кислоты, а оно, в свою очередь, - к повышению доли азота аминокислот в составе остаточного азота крови (гипераминоацидемии), общей гиперазотемии и аминоацидурии. Гипераминоацидемия, аминоацидурия и общая азотемия характерны для многих видов патологии А. о. При обширных поражениях печени и других состояниях, связанных с массивным распадом белка в организме, нарушаются процессы дезаминирования аминокислот и образования мочевины таким образом, что возрастают концентрация остаточного азота и содержание в нем азота аминокислот на фоне снижения относительного содержания в остаточном азоте азота мочевины (так называемая продукционная азотемия).
Продукционная азотемия, как правило, сопровождается выведением избытка аминокислот с мочой, поскольку даже в случае нормального функционирования почек фильтрация аминокислот в почечных клубочках происходит интенсивнее, чем их реабсорбция в канальцах. Заболевания почек, обтурация мочевых путей, нарушение почечного кровообращения приводят к развитию ретенционной азотемии, сопровождающейся нарастанием концентрации остаточного азота в крови за счет повышения содержания в крови мочевины (см. Почечная недостаточность ). Обширные раны, тяжелые и, инфекции, повреждения трубчатых костей, спинного и головного мозга, болезнь Иценко - Кушинга и многие другие тяжелые заболевания сопровождаются аминоацидурией. Она характерна и для патологических состояний, протекающих с нарушением процессов реабсорбции в почечных канальцах: болезни Вильсона - Коновалова (см. Гепатоцеребральная дистрофия ), нефронофтизе Фанкони (см. Рахитоподобные болезни ) и др. Эти болезни относятся к многочисленным генетически обусловленным нарушениям А. о. Избирательное нарушение реабсорбции цистина и цистинурия с генерализованным нарушением обмена цистина на фоне общей аминоацидурии сопровождает так называемый цистиноз. При этом заболевании кристаллы цистина откладываются в клетках ретикулоэндотелиальной системы. Наследственное заболевание фенилкетонурия характеризуется нарушением превращения фенилаланина в тирозин в результате генетически обусловленной недостаточности фермента фенилаланин - 4-гидроксилазы, что вызывает накопление в крови и моче непревращенного фенилаланина и продуктов его обмена - фенилпировиноградной и фенилуксусной кислот. Нарушение превращений этих соединений характерно и для вирусного гепатита.Тирозинемию, тирозинурию и тирозиноз отмечают при ах, диффузных заболеваниях соединительной ткани (коллагенозах) и других патологических состояниях. Они развиваются вследствие нарушения трансаминирования тирозина. Врожденная аномалия окислительных превращений тирозина лежит в основе алкаптонурии, при которой в моче накапливается непревращенный метаболит этой аминокислоты - гомогентизиновая кислота. Нарушения пигментного обмена при гипокортицизме (см. Надпочечники ) связаны с угнетением превращения тирозина в меланин вследствие ингибирования фермента тирозиназы (полное выпадение синтеза этого пигмента характерно для врожденной аномалии пигментации - а).
При массивном распаде клеточных структур (голодание, тяжелая мышечная работа, инфекции и др.) отмечают патологическое нарастание концентрации остаточного азота за счет увеличения относительного содержания в ней азота мочевой кислоты (в норме концентрация мочевой кислоты в крови не превышает - 0,4 ммоль/л ).
В пожилом возрасте снижаются интенсивность и объем синтеза белка за счет непосредственного угнетения биосинтетической функции организма и ослабления его способности усваивать аминокислоты пищи; развивается отрицательный азотистый баланс. Нарушения обмена пуринов у людей пожилого возраста приводят к накоплению и отложению в мышцах, суставах и хрящах солей мочевой кислоты - уратов. Коррекция нарушений А. о. в пожилом возрасте может быть осуществлена за счет специальных диет, содержащих полноценные животные белки, витамины и микроэлементы, с ограниченным содержанием пуринов.
Азотистый обмен у детей отличается рядом особенностей, в частности положительным азотистым балансом как необходимым условием роста. Интенсивность процессов А. о. на протяжении роста ребенка подвергается изменениям, особенно ярко выраженным у новорожденных и детей раннего возраста. В течение первых 3-х дней жизни азотистый баланс отрицателен, что объясняется недостаточным поступлением белка с пищей. В этот период обнаруживается транзиторное повышение концентрации остаточного азота в крови (так называемая физиологическая азотемия), иногда достигающее 70 ммоль/л ; к концу 2-й нед.
жизни концентрация остаточного азота снижается до уровня, отмечаемого у взрослых. Количество выделяемого почками азота нарастает в течение первых 3-х дней жизни, после чего снижается и вновь начинает увеличиваться со 2-й нед. жизни параллельно возрастающему количеству пищи.Наиболее высокая усвояемость азота в организме ребенка наблюдается у детей первых месяцев жизни. Азотистый баланс заметно приближается к равновесию в первые 3-6 мес. жизни, хотя и остается положительным. Интенсивность белкового обмена у детей достаточно высока - у детей 1-го года жизни обновляется около 0,9 г белка на 1 кг массы тела в сутки, в 1-3 года - 0,8 г/кг/ сут., у детей дошкольного и школьного возраста - 0,7 г/кг/ сут.
Средние величины потребности в незаменимых аминокислотах, по данным ФАО ВОЗ (1985), у детей в 6 раз больше, чем у взрослых (незаменимой аминокислотой для детей в возрасте до 3 мес. является цистин, а до 5 лет - и гистидин). Более активно, чем у взрослых, протекают у детей процессы трансаминирования аминокислот. Однако в первые дни жизни у новорожденных из-за относительно низкой активности некоторых ферментов отмечаются гипераминоацидемия и физиологическая аминоацидурия в результате функциональной незрелости почек. У недоношенных, кроме того, имеет место аминоацидурия перегрузочного типа, т.к. содержание свободных аминокислот в плазме их крови выше, чем у доношенных детей. На первой неделе жизни азот аминокислот составляет 3-4% общего азота мочи (по некоторым данным, до 10%), и лишь к концу 1-го года жизни его относительное содержание снижается до 1%. У детей 1-го года жизни выведение аминокислот в расчете на 1 кг массы тела достигает величин выведения их у взрослого человека, экскреция азота аминокислот, достигающая у новорожденных 10 мг/кг массы тела, на 2-м году жизни редко превышает 2 мг/кг массы тела. В моче новорожденных повышено (по сравнению с мочой взрослого человека) содержание таурина, треонина, серина, глицина, аланина, цистина, лейцина, тирозина, фенилаланина и лизина. В первые месяцы жизни в моче ребенка обнаруживаются также этаноламин и гомоцитруллин. В моче детей 1-го года жизни преобладают аминокислоты пролин и [гидр]оксипролин.
Исследования важнейших азотистых компонентов мочи у детей показали, что соотношение мочевой кислоты, мочевины и аммиака в процессе роста существенно изменяется. Так, первые 3 мес. жизни характеризуются наименьшим содержанием в моче мочевины (в 2-3 раза меньше, чем у взрослых) и наибольшей экскрецией мочевой кислоты. Дети в первые три месяца жизни выделяют 28,3 мг/кг массы тела мочевой кислоты, а взрослые - 8,7 мг/кг . Относительно высокая экскреция у детей первых месяцев жизни мочевой кислоты способствует иногда развитию мочекислого инфаркта почек. Количество мочевины в моче нарастает у детей в возрасте от 3 до 6 месяцев, а содержание мочевой кислоты в это время снижается. Содержание аммиака в моче детей в первые дни жизни невелико, но затем резко возрастает и держится на высоком уровне на протяжении всего 1-го года жизни.
Характерной особенностью А. о. у детей является физиологическая креатинурия. Креатин обнаруживается еще в амниотической жидкости; в моче он определяется в количествах, превышающих содержание креатина в моче взрослых, начиная с периода новорожденности и до периода полового созревания. Суточная экскреция креатинина (дегидроксилированного креатина) с возрастом увеличивается, в то же время по мере нарастания массы тела ребенка относительное содержание азота креатинина мочи снижается. Количество креатинина, выводимого с мочой за сутки, у доношенных новорожденных составляет 10-13 мг/кг , у недоношенных 3 мг/кг , у взрослых не превышает 30 мг/кг.
При выявлении в семье врожденного нарушения А. о. необходимо проведение медико-генетического консультирования .
Библиогр.: Березов Т.Т. и Коровкин Б.Ф. Биологическая химия, с. 431, М., 1982; Вельтищев Ю.Е. и др. Обмен веществ у детей, с. 53, М., 1983; Дудел Дж. и др. Физиология человека, пер. с англ., т. 1-4, М., 1985; Зилва Дж.Ф. и Пэннелл П.Р. Клиническая химия в диагностике и лечении, пер. с англ., с. 298, 398, М., 1988; Кон Р.М. и Рой К.С. Ранняя диагностика болезней обмена веществ, пер. с англ., с. 211, М., 1986; Лабораторные методы исследования в клинике, под ред. В.В. Меньшикова, с. 222, М., 1987; Ленинджер А. Основы биохимии, пер. с англ., т. 2, М., 1985; Мазурин А.В. и Воронцов И.М. Пропедевтика детских болезней, с. 322, М., 1985; Руководство по педиатрии, под. ред. У.Е. Бермана и В.К. Вогана, пер. с англ., кн. 2, с. 337, VI., 1987; Страйер Л. Биохимия, пер. с англ., т. 2, с. 233, М., 1985.